 |
A book by William H. Calvin UNIVERSITY OF WASHINGTON SEATTLE, WASHINGTON 98195-1800 USA |
|
THE CEREBRAL CODE Thinking a Thought in the Mosaics of the Mind Available from MIT Press in September 1996 copyright ©1996 by William H. Calvin |
|
Managing the Cerebral Commons
The map is not the territory, as various people said earlier in the century, but Arbib goes on to emphasize that our job as brain theorists is not to replace a “life richly lived by the running of some computer program.” We’ve got to extract something simpler, a maplike guide to the underlying complexity, despite the seeming functional incomprehensibility of those cells in the hidden layers in connectionist networks. We need brain abstractions — such as common processes, used again and again — in order to appreciate the fine details. Among the abstractions that have recently been helpful are those six essentials of a darwinian process: a characteristic pattern that is copied, occasional variants that compete for a work space, with success biased by a multifaceted environment, and with the more successful producing most of the next round of slight variants (Darwin’s inheritance principle). In the last chapter, we saw how copying mechanisms also helped identify a characteristic pattern. The predicted mosaics of electrical activity are just one ephemeral state of the cerebral cortex, but they may provide a guide to what else it does. If we can learn to think about the dynamically reforming patchwork quilt of cloning territories as simply as we think about maps, perhaps we can find a way around the hidden-layer incomprehensibility. This chapter (far easier than the last two) will address the variant patterns and the work space itself, and later chapters will consider cloning competitions in more detail. In between is some chaos, in the best sense of the word, that underlies the memorized environment. But first we are going to need some metaphors to guide us in exploring a jack-of-all-trades work space. We need a lot of multifunctional potential (potentially powerful, but it makes it difficult to name things) rather than a lot of fixed compartments.
|
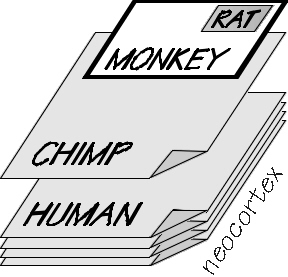 We humans have a lot of cerebral cortex,
enough to cover four sheets of typing paper;
that’s the potential size of our patchwork quilt
of competing hexagonal patterns. A chimpanzee’s cerebral cortex is equally thick but there’s
only enough, when all is flattened out, to cover one sheet of paper. Monkeys have about a
postcard worth, and rats are down in postage stamp territory. So far, after a century of
neuroscience, there’s nothing to suggest that a square millimeter worth of human cortex is much
different than the same area in a monkey’s brain.
We humans have a lot of cerebral cortex,
enough to cover four sheets of typing paper;
that’s the potential size of our patchwork quilt
of competing hexagonal patterns. A chimpanzee’s cerebral cortex is equally thick but there’s
only enough, when all is flattened out, to cover one sheet of paper. Monkeys have about a
postcard worth, and rats are down in postage stamp territory. So far, after a century of
neuroscience, there’s nothing to suggest that a square millimeter worth of human cortex is much
different than the same area in a monkey’s brain.But, though everyone assumes that bigger is likely better, we’ve had trouble saying in what way that four times as much cortex has been better. There are increases everywhere in the forebrain structures during the evolution from apes to humans. Yes, there are some human abilities such as language, music, planning ahead, accurate throwing, and toolmaking that might require some additional cortical space beyond that available in the apes, but they haven’t been “tacked on” as a bulge here and a bump there, in the way a house is often enlarged over the years. It seems to be easier to increase all of cortex than just to selectively enlarge one piece. This wouldn’t have surprised Charles Darwin, who emphasized specialized adaptations but also acknowledged that the underlying variations often involved quite general aspects of growth and form, inextricably linking some functions. Many of the new or improved-beyond-the-apes human skills seem to share cortical space with one another, allowing for conversions of function (as I discuss in How Brains Think, conversion and coexistence of functions in the same structure is yet another original insight of Darwin’s). And such multifunctionality has some interesting implications for the evolution of new functionality. You wouldn’t think that a new function could spring forth fully formed, as Minerva leaped forth from the head of Jupiter — but it can, thanks to sharing space with an older function. Such bundling of functions constitutes something of a “free lunch.” The new function might profit from some additional improvements for efficiency, but its invention per se is not likely to be understood by concentrating on such slow adaptations.
|
|
New functions usually get a big head start, making part-time use of an organ that, in the
past, has been under natural selection for some other, and often quite different, function.
Feathers for thermal insulation serving to improve wing aerodynamics is a standard example.
Just as the 1940s computers for missile guidance gave a big boost to computers for word
processing a few decades later, so brains are particularly capable of conversions of function,
improvements in one function stage-setting the emergence of other, seemingly unrelated
functions in the off-hours for the original function. The functions might then continue to share
the same space, or the space might subdivide into specialists in the two functions. Because of co-habitation of functions, the labels we use can create some serious blinders. For example, large areas of “language cortex” may be involved with both hand and face movement sequencing — and with analyzing the sequence of sounds you hear. Language cortex isn’t exclusively concerned with language. Sensory and motor, hand and face, illustrate the multifunctional reality that brain theories need to cover.
In the experiments of generations of neurophysiologists, individual cortical neurons have often responded to several sensory modalities. Neuroanatomists know that everything is connected to everything (if not directly, then within only several removes). Where they exist, specializations are surely not exclusive, probably in much the same sense that an underutilized neurosurgeon could on occasion help out by doing routine life-insurance physicals in a general medical practice.
|
 Certainly, just reading the popular press accounts about advances in neuroscience, you’d think
that cortex was very specialized, with pigeonholes everywhere: areas for inanimate objects,
others for tool concepts, and so on. This recalls Cicero’s advice about how to remember new
names, which stressed imagining each new person in a different location, such as a room within
a large mansion (offering similar advice thirteen centuries later, Thomas Aquinas suggested
niches in a cathedral). But, however convenient it may be initially to think of the brain as a
tabula rasa, with empty pigeonholes at birth waiting to be filled up with a lifetime of memories
and acquired skills, few neuroscientists think that it really works that way. Nevertheless, we
happily analyze whatever specialties we can find — and, by the functional names we give the
area, often blind ourselves to considering additional coexisting functions.
Certainly, just reading the popular press accounts about advances in neuroscience, you’d think
that cortex was very specialized, with pigeonholes everywhere: areas for inanimate objects,
others for tool concepts, and so on. This recalls Cicero’s advice about how to remember new
names, which stressed imagining each new person in a different location, such as a room within
a large mansion (offering similar advice thirteen centuries later, Thomas Aquinas suggested
niches in a cathedral). But, however convenient it may be initially to think of the brain as a
tabula rasa, with empty pigeonholes at birth waiting to be filled up with a lifetime of memories
and acquired skills, few neuroscientists think that it really works that way. Nevertheless, we
happily analyze whatever specialties we can find — and, by the functional names we give the
area, often blind ourselves to considering additional coexisting functions.|
So the specialized-room analogies won’t do, in the long run, however useful they may be for
analyzing early stages of perception. I’m going to introduce multifunctionality via some fanciful
allusions to the various fields and alcoves within a large public park. Grassy parks such as
London’s Hampstead Heath are nice, but I have been intrigued with the grassless parks in some
cities of Europe, where fine brown gravel walkways cover everything that isn’t fenced off. The park fences are probably due to another tragedy of the commons, I thought, a consequence of the increasing population density inside cities. With more people over the centuries but even less parkland, the open space becomes overused. And then come the fences. Public spaces such as communal pasture lands have some drawbacks that fencing may avoid, such as overgrazing (where the consequences accrue to the owner of the fenced area, rather than being spread out over everyone who shares the commons). That might, I thought, be one advantage of the departmentalization of the cortex, all those Brodmann Areas. Multifunctional areas of cerebral cortex may, of course, have all the problems associated with a commons — and the tragedies thereof. Indeed, I observed from a park bench in the center of Paris, some of the graveled areas are flattened so evenly that — well, you don’t suppose that the workers have been preparing the surface for the next obvious step?
|
 The schoolchildren who play in the park will first create a hexagonal mold, its bottom carved into a mascot design. Then they’ll pour and lay as many of them as they can, design side up. Some areas will have a standard cat design, and then you might walk across some gravel again until encountering another honeycombed area, each paver having the dog design favored by another group of paver-layers. Since they’re all hexagonal in shape, they’re easy for the children to lay without exercising any judgment, a nice algorithmic process. A robot could do it.
What, you wonder, will happen when the army of cat pavers comes face to face (well, edge to edge) with the dog ones? Will dog pavers replace cat pavers and start taking over the cat territory? Is the size of the dog territory important for some reason? Now, if these were the hexagonal mosaics predicted for cerebral cortex, someone would surely suggest that they have to all fire in synchrony, binding them together so that they will be noticed by an observer on high. I’d prefer to suggest (just wait for chapter 8) that there’s a critical number needed before they’re likely to communicate effectively with the motor centers of the brain. Indeed (aren’t reveries fun?), leaving aside the cartesian theater fallacy for the moment, what might you see from an aerial photo of this park? Probably lots of alcoves (thanks to those barriers known as foliage) opening off the central spaces, though the extent of the alcoves will be obscured by the overhanging tree branches, as will some of the paths between them (that’s one advantage of a map over an aerial photo — and shows why an abstract schema can improve on a “rich mental image”).
|
 Perhaps, I thought, we could
even dispense with the paver-layers and imagine a pair of
pavers as sometimes cloning a
third with the same characteristic
decoration, inserting it into any
adjacent unpaved space.
Perhaps, I thought, we could
even dispense with the paver-layers and imagine a pair of
pavers as sometimes cloning a
third with the same characteristic
decoration, inserting it into any
adjacent unpaved space. Up close, of course, there are many things going on
simultaneously in this park: wandering around it
might reveal country dancing in one alcove, children
listening to a storyteller in another alcove,
occasionally an alcove with some specialized facilities
such as chess tables or a few charcoal grills. Though
not suitable for the dancers, such specialized facilities
could still be used by the storyteller. Various paver
designs might get started in one alcove or another,
with themes reflecting the activities in progress, and
then clone their way outward into more general-purpose open spaces, the ones where soccer games
alternate with rock concerts and frisbee tag.
Up close, of course, there are many things going on
simultaneously in this park: wandering around it
might reveal country dancing in one alcove, children
listening to a storyteller in another alcove,
occasionally an alcove with some specialized facilities
such as chess tables or a few charcoal grills. Though
not suitable for the dancers, such specialized facilities
could still be used by the storyteller. Various paver
designs might get started in one alcove or another,
with themes reflecting the activities in progress, and
then clone their way outward into more general-purpose open spaces, the ones where soccer games
alternate with rock concerts and frisbee tag.|
I’ve long thought that copying competitions,
rather like the paving of that park, must be the way in
which my premotor cortex goes about deciding
between various alternative actions as I make up my
mind. Thumb-up, a precision grip, and pointing might be the contending possibilities. A
hexagonal movement schema is, in principle, no different than a sensory one. To an observing
neurophysiologist, both would likely be as incomprehensible in appearance as a bar code on a
grocery shelf.
I expect that for well-learned “cortical reflexes,” as when I quickly wave at a friend driving past, the decision making is much simpler. But when there is time for indecision, and where there are various possible actions contending, a copying competition is a nice mechanism for gating thought into action, one solution to the problem of how billions of neurons can make up their collective mind. Perhaps it’s a matter of whose song can recruit the largest chorus, and not a manner of annexing all the competing territories — rather as in hegemony, where one nation may dominate the political agendas of a region without actual territorial conquest.
|
 Perhaps there is some
minimum plurality needed to
get the action underway.
Perhaps it takes a certain
amount of coherent cortical
output, enough hexagons
worth of movement schemas
singing in sync as a chorus, in
order to persuade the
subcortical centers to get their
act together.
Perhaps there is some
minimum plurality needed to
get the action underway.
Perhaps it takes a certain
amount of coherent cortical
output, enough hexagons
worth of movement schemas
singing in sync as a chorus, in
order to persuade the
subcortical centers to get their
act together.|
We have already identified the first two darwinian essentials, and now we have a candidate
for a work space. It’s just an analogy so far, but soon it will be fleshed out as like a dynamically
changing patchwork quilt of differing hexagonal territories. What about variations on the
cloned pattern, ways of making a new individual, one that might clone a new patch and
compete with its parent pattern? There is clearly the possibility of the aforementioned “error correction” preventing any variations on an established pattern, once a given hexagon is surrounded by like-minded ones for crystallization tendencies to assert themselves. But on the edges of a paver area, bordering an unorganized area, a hexagon may only have three or four neighbors. Furthermore, some geometric arrangements of barriers allow only two neighbors to an unorganized hexagon. It is on such frontiers that we should first look for variant patterns, not within the “continents.” Most simply, a hexagonal variant could be caused by the failure of one or more of the triangular arrays to clone new nodes. The spatiotemporal melody within the hexagon would be missing a note, in the manner of a piano with a dead key. This “loss of detail” might mean the dropout of one attribute, such as color, but not necessarily. When we say yellow or horizontal, we are only emphasizing the most easily investigated attribute of our “feature detector” (itself a convenient fiction). Many interneurons are multisensory and so a given triangular array need not represent a modality that is orthogonal to all other attributes.
|
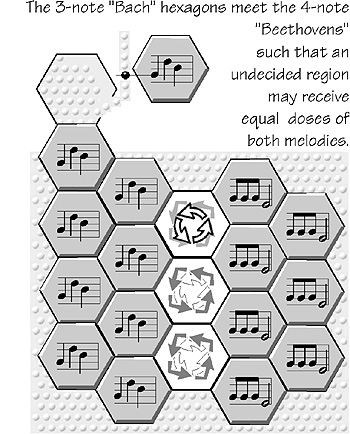 A second type of variant would be when new
triangular arrays join the collection. The most likely
way for this to happen is when two different
hexagonal mosaic patterns collide and a “no-man’s-land” forms where the two spatiotemporal patterns
superimpose (or perhaps they partially inhibit one
another, in the clash of incompatible spatiotemporal
melodies). There are two aspects of such a no-mans-land: variant patterns and whether the border moves
as one tiling replaces another (to be treated in the next
chapter, after we examine resonances).
A second type of variant would be when new
triangular arrays join the collection. The most likely
way for this to happen is when two different
hexagonal mosaic patterns collide and a “no-man’s-land” forms where the two spatiotemporal patterns
superimpose (or perhaps they partially inhibit one
another, in the clash of incompatible spatiotemporal
melodies). There are two aspects of such a no-mans-land: variant patterns and whether the border moves
as one tiling replaces another (to be treated in the next
chapter, after we examine resonances).|
An evolutionary work space is not usually a
continent but rather a much smaller region, such as an
island or isolated valley or hilltop. Patchy distribution
of essential resources can create subpopulations, if
migration across the resourceless gaps is infrequent.
I’ll develop the idea further in Act II, but a work space
implies more than a blackboard; it’s also a region
where a subpopulation can go extinct, and thereby create an empty niche. So, too, the relevant cortical work space for cloning competitions is unlikely to be the whole neocortex. The change in the metric associated with the gaps (and the basal dendritic spreads) between adjacent cortical areas may ensure that cloning remains within one Brodmann Area (though the U-fibers of the better-known corticocortical connections might still manage to produce clones, as I will show in chapter 8). Patchy excitatory drive may create even smaller work spaces, as might waves of inhibition. Much more can (and will) be said about the generation of variants and the size of work spaces, but it will be useful to first consider some cortical possibilities for that fifth essential, the multifaceted environment, now that we have identified the pattern, how it copies, how it might vary, and the limited territory in which it might compete.
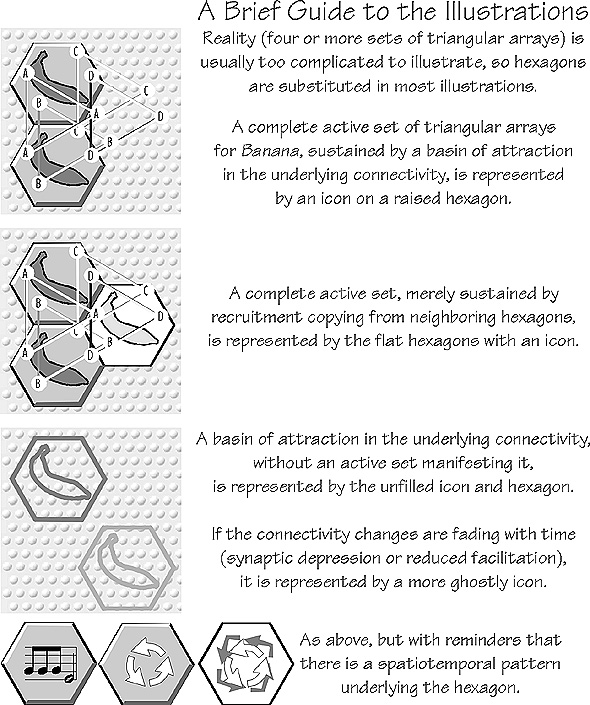
|
 You are reading THE CEREBRAL CODE.
You are reading THE CEREBRAL CODE.
The paperback US edition |