[What] you are describing is not an object but a function, a role that is
inextricably tied to some context. Take away the context, and the meaning also
disappears.... When you perceive intelligently, as you sometimes do, you always
perceive a function, never an object in the...physical sense.... Your Cartesian idea
of a device in the brain that does the registering is based on a misleading analogy
between vision and photography. Cameras always register objects, but human
perception is always the perception of functional roles. The two processes could
not be more different....
the mathematician Stanislaw Ulam, about 1970
IT IS CURIOUS HOW DISTRACTING little black lines in the sky prove to be, as if they had
some privileged access to our brains -- like visual equivalents of advertising jingles hung from
utility poles. And they do have priviledged access, another discovery that Steve Kuffler had a
hand in.
Leaving the Church of the Messiah, one walks east along a winding road in Woods Hole that
would be beautiful except for the proliferating poles stringing the skyline with metastasizing
threads of metallic tumor. It's an area of expensive homes whose owners could afford to pay for
burying the utility wires, but only people get buried around here. In another block down Church
Street, the wires thin out and the view opens onto Vineyard Sound, a long, sandy beach
stretching into the distance. At the end of the beach, a white pillar rises into the blue sky, capped
with black. It is surely the Nobska Point lighthouse, and it looks somewhat familiar.
A lighthouse is a version of the scarecrow, but meant to warn off sailors. And just as the
scarecrow is a human mimic of sorts, so the lighthouse stands as a surrogate human, erected to
send a human message in times of need. As lighthouses get "smarter" with the addition of
automatic fog sensors that trigger their foghorns, they come to be more and more like a stationary
robot. Their foresightful designers have endowed the lighthouse with ways of forecasting
trouble, and appropriately responding. Lighthouses lack the versatility to be considered
conscious, but the air-traffic control system might someday evolve into a partially conscious
robot, constantly trying out "what if" scenarios and so heading off collisions between airplanes in
much the same ways as human controllers now attempt to do.
WALKING UP THE BEACH IN THE WET SAND just above the lapping waves of Vineyard
Sound, I saw various people from the MBL and WHOI, talking science. Bankers from Boston
can also be found on this beach, though they do not draw diagrams of cells in the sand, so far as I
know. The diagrams may be washed away by the next high tide, but the new concepts linger on.
This beach has seen a lot of science happen, as has Stony Beach near MBL, which Lewis Thomas
immortalized in his essay "The MBL," in Lives of a Cell:
It is so crowded that one must pick one's way on tiptoe to find a hunching place, but there is
always a lot of standing up anyway; biologists seem to prefer standing on beaches, talking at each
other, gesturing to indicate the way things are assembled, bending down to draw diagrams in the
sand. By the end of the day, the sand is crisscrossed with a mesh of ordinates, abscissas, curves
to account for everything in nature.
You can hear the sound from the beach at a distance, before you see the people. It is that most
extraordinary noise, half-shout, half-song, made by confluent, simultaneously raised human
voices, explaining things to each other.
This beach on Vineyard Sound is instead spread over two blocks and has a higher proportion of
sand castles than graphs. The lighthouse down at the end of the beach, as one walks closer and
the perspective changes, evokes ever stronger sensations of déjà vu -- I seem to have some
mental image that this picture is triggering.
And then I remember the art museum where I've seen it before: it's a close relative of the
lighthouse that Edward Hopper painted. And painted again and again, though they might be
different lighthouses, because the coast guard built a number of them from the same 1870's
design; they can be found all around Cape Cod and the offshore islands. This lighthouse looks
even more solid than the painted versions, ready to take whatever the weather dishes out next.
I HAVE AN EDWARD HOPPER SCHEMA somewhere in my head, not to mention a Picasso
schema, and a Henry Moore schema, and so on. A schema is a mental image. While schema
was initially used to avoid using memory trace (another term invented to try to pin down an
elusive concept), a schema is not just any memory. A sensory schema is a schematic outline that
fits most of the likely variations that Hopper constructed; it fits in a fancier way than a cookie-cutter fits a Christmas cookie, but that's the general idea.
A collection of schemas -- say, my collection of mental images of artistic styles -- is something
like a family of cookie-cutters, each shape being tried out on a particular Christmas cookie to see
which comes closest. The collection of schemas seems to sit there in your head, constantly on
the lookout for something that fits it. So that when you see a fragmentary Picasso sketch, and
something shouts "Picasso!" inside your head, you've heard a schema speaking.
I also have a lighthouse schema, equally well activated by English lighthouses and Hawaiian
lighthouses. It is only the lighthouses that are in the local 1870's style that additionally activate
my "Edward Hopper" schema, and then only when seen from a perspective similar to the ones
that Hopper favored (aerial photographs won't do). Such an "Edward Hopper schema" is a
collective memory, containing something of the "essential" features of his style and subject
matter that come through whatever variations are present in a particular example. For every
word in your vocabulary, you probably have one or more schemas -- though I certainly have
some schemas for which there is no word in my vocabulary, such as people that I've seen before
but whose names I haven't learned yet. And some schemas aren't objects but movements: When
we construct a sentence or spin a scenario, the units that we are chaining together are primarily
sensory ("noun") and movement ("verb") schemas. You, me, food, rock -- as well as run, walk,
gallop, shuffle, touch, bend, break.
EPICURUS AND LUCRETIUS considered mental images to be simulacra, and Aristotle
compared them to "the imprint left by a seal on a wax tablet." They hadn't seen cookie-cutters.
And now we have some even better analogies -- indeed, potential mechanisms. The lighthouse
beacon winking on and off (even in broad daylight, the 150-watt light bulb looks bright because
of that old 1825 Fresnel lens that focuses its photons) reminds me of another Steve Kuffler
classic. His 1953 work was on how nerve cells in the eye respond to such lights winking on and
off; I'm unlikely ever to forget it, as reading it about six years after it was published was one of
the major reasons that I had forsaken physics for physiology. It is still a classic, required reading
for the aspiring graduate student. It demonstrates a cellular version of the schema -- or at least
one of its building blocks.
The story starts even earlier, in 1938, when H. K. Hartline studied frog retinas and their response
to winking lights. He found that each optic-nerve "wire" responded to far more than just one
spot on the retina -- it was a whole patch (which he called the cell's "receptive field"). It would
fire a train of spikes in a staccato manner whenever the light was turned on, but then settle down,
not acting much different than in darkness. But when the light was finally turned off, it gave
another burst of activity, a cellular version of "Hey! Who turned the lights off?" This OFF-response was a puzzle, though not unrelated to the problem of feeling your wristwatch's absence
just after you remove it from your wrist and all those flattened hairs start popping back up.
A friendly caution: You are approaching some conceptually difficult material in
the remainder of this chapter, and at places in the following two chapters.
Ordinary mortals will likely, at some point, feel disoriented -- and for good reason.  Everyone who has learned this material has complained too. Graduate and
medical students traditionally stumble on "receptive fields" -- but then, like art
students who finally learn how to "see" a particular painting, recover and soon
sort out a mental picture of how a brain cell sees the world.
The "viewpoint" turns out to be all-important, both the cell's viewpoint and the
scientist's varied viewpoints used to analyze the cell's properties. We're still
debating how a committee of cells, like a jury of critics, can have a collective
viewpoint!
None of this material is essential, in that you could skip over it and still understand
the rest of the book, but I include it because so many of those who come to
understand it are, let us say, inordinately impressed. It provides a nice, concrete
example (almost too nice!) of how the brain goes about its business. Indeed, to
those familiar with business accounting, the time history of the cell (such as those
OFF-responses) is like cash flow tracking, and the Mexican hat "receptive field"
represents another way of looking at much the same data, showing where the
income and expenses originated in a given time period and how much profit was
paid out to shareholders. Anyone familiar with accounting practices may have a
better background for appreciating this material than the typical neurobiology
graduate student.
Everyone who has learned this material has complained too. Graduate and
medical students traditionally stumble on "receptive fields" -- but then, like art
students who finally learn how to "see" a particular painting, recover and soon
sort out a mental picture of how a brain cell sees the world.
The "viewpoint" turns out to be all-important, both the cell's viewpoint and the
scientist's varied viewpoints used to analyze the cell's properties. We're still
debating how a committee of cells, like a jury of critics, can have a collective
viewpoint!
None of this material is essential, in that you could skip over it and still understand
the rest of the book, but I include it because so many of those who come to
understand it are, let us say, inordinately impressed. It provides a nice, concrete
example (almost too nice!) of how the brain goes about its business. Indeed, to
those familiar with business accounting, the time history of the cell (such as those
OFF-responses) is like cash flow tracking, and the Mexican hat "receptive field"
represents another way of looking at much the same data, showing where the
income and expenses originated in a given time period and how much profit was
paid out to shareholders. Anyone familiar with accounting practices may have a
better background for appreciating this material than the typical neurobiology
graduate student.
This "Somebody turned the lights off" report (an OFF-response) is an exaggerated aspect of
"temporal contrast," a sensitivity to changes in light level rather than absolute light level. It is
why flashing or moving lights grab your attention better than steady lights. Keffer Hartline
(another luminary of MBL) also noted in passing that the background was important in
influencing the vigor of these staccato responses, rather as the brightness of the sky behind the
lighthouse affects how bright the beacon seems. Robert Barlow (once Hartline's graduate
student, and now a veteran of the Woods Hole science scene) noted this spatial-contrast effect in
1953, as did Stephen Kuffler. Temporal contrast and spatial contrast turn out to be the major
building blocks of visual perception, with color contrast an important added feature in some
animals such as the primates. Our ancestors made their living by being able to spot ripening fruit
high up in the trees against a background of waving leaves; that green spot you sometimes see
from the Nobska lighthouse alongside the setting sun is one of the collateral consequences.
Kuffler dug deeper and wound up showing the underlying mechanism of spatial contrast: each of
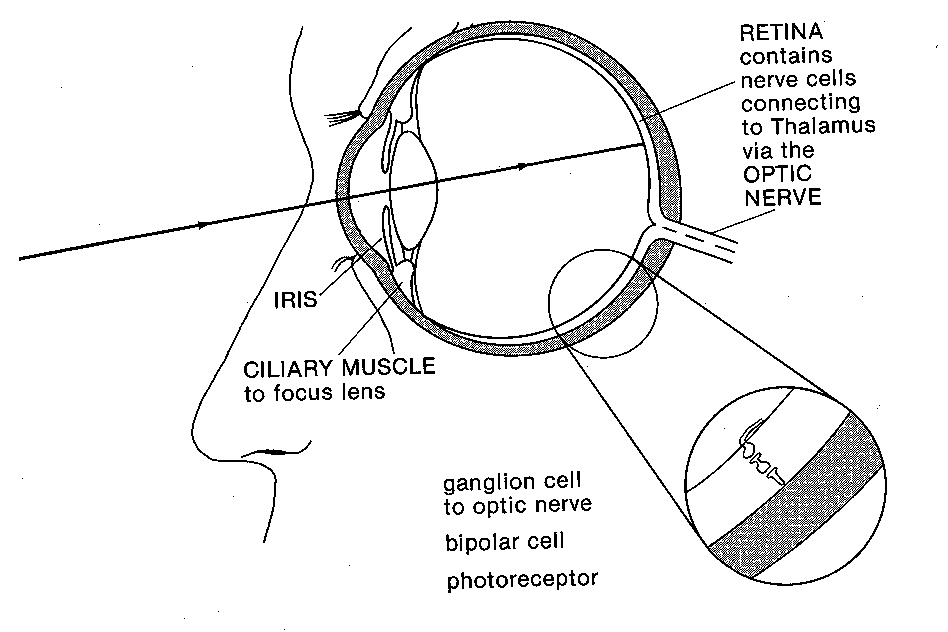
the cat's retinal ganglion cells (which are typically third in the chain of cells between
photoreceptor and brain) was receiving input from thousands of photoreceptors (via intermediate
cells called bipolar cells and amacrine cells), rather as a funnel collects raindrops from a wide
area and concentrates it into a narrow stream. There has to be some funneling, as there are about
a hundred photoreceptors for each "wire" back to the brain (the retinal ganglion cell axon; about
a million of them constitute an optic nerve). And it isn't a matter of a hundred photoreceptors
connecting to each ganglion cell: Messages from thousands of photoreceptors are funneled into
each ganglion cell, except some cancel out the actions of others.
Each of those little "wires" in the optic nerve sends messages akin to a bank statement when it
tells you how much interest was paid this month. You have to imagine, instead of an eye, a giant
bank that is busily mailing out a million statements every second. What maximizes the payout
on a single wire? That depends on the bank's rules, and how you play the game.
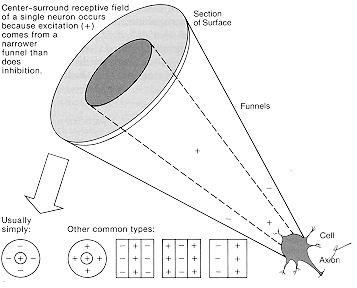
The plus-and-minus, push-and-pull arrangement (which neurophysiologists tend to call
"excitation" and "inhibition" instead) is not unlike the deposits to and withdrawals from my bank
account. The output (the interest paid out) is naturally proportional to the net balance. There is
often a minimum requirement ("5 percent interest paid on all balances over $1,000"). Nerve
cells don't compound interest (since they pay it out each time) -- but they have other nice features
that bank accounts lack. I sure wish that my bank account paid an "adaptation bonus" whenever
I resumed depositing money after a fallow period! Some nerve cells even have "OFF-responses"
as a rebound from inhibition, equivalent to a bank paying a bonus when you stop a long series of
withdrawals. I must remember to suggest that to my bank; they can advertise it as "the brain's
way of doing business."
Kuffler discovered the parts of the retinal mosaic from which "deposits" and "withdrawals" each
originated, something that Hartline couldn't resolve in frogs. A single retinal ganglion cell really
receives two funnels, a wide funnel and a more potent narrow funnel; one inhibits and the other
excites the retinal ganglion cell. It's not unlike the sources and sinks of a checking account: We
usually funnel into our account a small number of large deposits, and write a much larger number
of small-amount checks.
A uniform illumination of the back of the eye, such as one gets from looking at the blue sky,
stimulates both funnels, and the messages cancel much of the time. A small spot of light such as
the lighthouse beacon might fill the small funnel but only half fill the larger one. If the large but
weak funnel was connected with inhibition, and the small but potent funnel with excitation, the
net difference would cause a vigorous response. Fill up more of the large but weak inhibitory
funnel by using a larger spot of light, and the excess of excitation over inhibition would decline
and the staccato response would decrease to a whimper.

The overall result after subtraction looks something like a doughnut, with a center and a
surround. An even better analogy is the Mexican hat, peaked in the middle and flanked by wide
shallow troughs: That's the size of the interest paid out for a small spot of light at each position
in space. When you adapt to darkness (your night vision improves after about twenty minutes in
the dark), that wide inhibitory funnel is disconnected from the retinal ganglion cell as a way of
"running wide open", getting maximum sensitivity by not subtracting anything. Again I wish
that my bank would let me boost my interest payments by simply not writing any checks for a
few months. Disconnecting the inhibitory funnel in dim light doesn't similarly result in big
problems later, unless you count the momentary blindness that occurs when you turn on the
bathroom light in the middle of the night (bathroom lights were not a big problem as humans
evolved during the ice ages).
So such a cell is a specialist in small spots of light -- indeed, the diameter of the optimal spot is
about the size of the hatband in that Mexican hat analogy. Such a cell discriminates against
larger spots, hat-brim width or more, sometimes refusing to tell the brain about them at all. But
that's all right, as there are other nearby cells specializing in big spots, having a somewhat
different arrangement of funnel sizes, just as there are hats with narrow brims for fat heads. We
tend to start thinking about such cells as specialists like cookie-cutters of a certain size, just
sitting there trying out each new spot of light to see if it's the right size. But the Mexican hat
arrangement is the better analogy, as it tells you what would happen to spots that weren't quite
the right size.
Now some bank accounts don't have the few-deposits-but-many-checks characteristics of my
checking account; they're more like a small business that receives many mail orders and has only
a few suppliers to pay. Some retinal cells are like that, with their funnels hooked up just the
opposite of the previous examples. Their weak but wide funnel is excitatory, and their narrow
but potent funnel is inhibitory. And so they seem to specialize in black spots on white
backgrounds rather than white spots on darker backgrounds; they're better at detecting the insects
on the sand beach (and the black periods ending sentences) than the white spots such as
lighthouse beacons against the night sky. This yields an inverted Mexican hat curve: A black
spot imaged onto the inner of the two-funnel arrangement doesn't cause any inhibition, and so the
excitatory response from the white background imaged onto the larger funnel has nothing to
balance it. Thus one gets a vigorous staccato burst to a black spot on a lighter background.
The map (such as the Mexican hat) of what's connected to a single cell is what is now called the
"receptive field" of that cell -- but one can also think of it as a template, the cell always being on
the lookout for images that fit its favored template. Just as cookie-cutters the size of the smaller
funnel won't fit a larger cookie, so receptive-field centers (the hat-band size) tend to represent the
optimal size of a stimulus spot. Specialists in small black spots on lighter backgrounds are
sometimes called "bug detectors," because they spring into activity when the image of a fly is
presented to them. But they also respond pretty well to thin black lines and usually to edges that
cover only half of a funnel. So they are not exclusively "bug detectors": The moving black bug
is simply their optimal stimulus. If you think of the bug as the "essence" of the cell's function,
you miss out on the whole range of less optimal, but still effective, stimuli that also run the cell.
In 1959 it was reported by Maturana, Lettvin, Pitts, and McCulloch that frogs were even more
discriminating. Their retinal ganglion cells didn't bother telling the frog's brain about anything
that wasn't moving food like flies, or a tilting horizon, or some other key feature of their
environment. Presented with a novel visual feature like a house or car, they often didn't bother
mentioning it at all to the brain (unless, of course, it was moving quickly toward them, something
that every visual system manages to sense). The frog's retina seemed to be a special-purpose
computer, not a TV camera that indiscriminately reports everything it sees. That was troubling:
Why was the frog throwing away all that more detailed information?
Doesn't he realize that he'll need it to read with, when he "grows up" evolutionarily? This is, of
course, our point of view; the frog is interested in more immediate things, like catching flies for
dinner, and has apparently tuned up his eye to maximize the detection of little moving black
spots. Our point of view might seem pretty parochial to the frog too. It would have a hard time
understanding our reductionist preoccupation with taking things apart into ever-smaller pieces:
discriminating between the fly's wings and feet might seem a little excessive to the pragmatic
frog.
Much of today's public anxiety about science is the apprehension that we may be
overlooking the whole by an endless, obsessive preoccupation with the parts.
the American physician and essayist Lewis Thomas
Behind the holists' very proper concern with the need to consider the system as a
whole lies also a fear that reductionism attempts not to just explain but to explain
away; that by reducing humans to an assemblage of working parts, their
humanity has been in some measure also reduced. This is an important fear,
and one which should be treated with respect.
the British neurobiologist Steven P. R. Rose
SPRINGY SAND? Walking on the sand beach on the eastern side of the lighthouse can be like
walking on a forest floor carpeted with pine needles where the floor gives and rebounds a little.
But here on this sandy beach one positively bounces. Why?
The "underlying mechanism" is not hard to discover, since it is literally underlying and can be
uncovered by digging down a little with one's toe: Great masses of seaweed have been washed
ashore during a recent storm, some stuffed with little air bladders that keep them floating up near
the light just under the ocean's surface. Then a windstorm washed sand up the beach to cover up
these mats of cushioned seaweed. And so one bounces along the sand like some improbable
ballet dancer.
Eventually, the seaweed cooks beneath the hot sand. Vegetation that gets buried deeper can turn
into oil; our cars are propelled on the cooked vegetation of great tropical swamps that got
themselves buried many millions of years ago. That some of the great oil fields are now located
up above the Arctic Circle shows you how much the continents drift around, as all that oil was
originally buried when it was within about 20° of the equator.
This also shows you what scientists mean by "reductionist" approaches: Springy sand attracts a
scientist who wants to know what the resiliency is all about. Its underlying mechanism,
discovered by "digging deeper," is buried seaweed. Then the agenda shifts: Why is the seaweed
bouncy? Its underlying mechanism is little air bladders. And why is that? It's due to the way
light is filtered by water, making things dark in the ocean depths: If you want to photosynthesize,
you want to stay as close to the surface as possible. And hence built-in flotation devices. Why
photosynthesize...? And so forth, with a constantly shifting agenda that maintains little of the
original question about resiliency (which, after all, was answered right off).
There are a lot of scientists who started out hoping to understand the mind who haven't been able
to answer such questions right off, and so have gotten distracted by underlying mechanisms and
the subtle meandering of the agenda. The only way to understand something thoroughly is to
investigate all its parts, but sometimes that isn't enough: Sometimes you have to figure out how
they work in committees, as when they regulate your body temperature or blood pressure to
remain within a narrow range of desirable levels. And so "understanding how we see something"
may involve a lot of analysis of the parts, but also figuring out how new properties emerge from
combinations.
Understanding emergents is generally harder than understanding the pieces, and an imperative of
science (or at least of scientists continually faced with raising money and training graduate
students) is to "make progress" via asking answerable questions. And some of our most
respected scientists are the ones who switch from subject to subject, constantly exploiting the
new techniques for "looking deeper." Only if they are disabled from laboratory work by illness
or retirement do they ever get around to "trying to piece things together." Yet the history of
science is written by both types, the experimenters like Hooke, Franklin, Boyle, and Faraday, and
the great synthesizers like Newton, Mendeleyeff, Einstein (Neils Bohr had to think for years
before coming up with the model of the atom that proved so successful). In eighteenth- and
nineteenth-century biology, the great synthesizers were even more prominent: Linnaeas,
Lamarck, Darwin. Neurobiology is young, and its history is mostly the history of the meandering
reductionists.
The important thing in science is not so much to obtain new facts as to discover
new ways of thinking about them.
the English physicist William Lawrence Bragg (1890-1971)
Many students of animal behavior have become so fascinated with its
directedness, with the question "What for?" or "Toward what end?" that they
have quite forgotten to ask about its causal explanation. Yet the great question...
"How?" [is] quite as fascinating as the question "What for?" -- only they
fascinate a different kind of scientist. If wonder at the directedness of life is
typical of the field student of nature, the quest for understanding of causation is
typical of the laboratory worker. It is a regrettable symptom of the limitations
inherent to the human mind that very few scientists are able to keep both
questions in mind simultaneously.
the Austrian ethologist Konrad Lorenz, 1960
VISUAL PERCEPTION MECHANISMS had been a mystery before all the wire-tapping
neurophysiology on frogs, cats, and monkeys; optics had yielded to investigation, and some
shrewd guesses (particularly Thomas Young's 1802 trichromaticity theory) had been made about
underlying color vision mechanisms, but no one had shown how the brain went about taking
apart an image into its component parts.
Now we had an impressive explanation of how the nervous system went about that, breaking the
image into components via how the excitation and inhibition were wired up. And a flurry of new
research started, analyzing the next stages of analysis back in the brain proper. We were
particularly interested in a visual area at the back of the head, which is where those messages in
the optic nerve seemed to be addressed. If that area, called "primary visual cortex" or "Area 17"
or sometimes just "V1," wasn't working because of damage, people seemed to be functionally
blind.
Two of Steve Kuffler's immediate followers were the young ophthalmologists-turned-neurophysiologists, David Hubel and Torsten Wiesel. When Kuffler went to Harvard Medical
School in the late fifties, he hired the pair -- and then went back to studying his first love,
synapses (in 1961, together with Josef Dudel, he discovered a major new principle we call
presynaptic inhibition, which results in a percentage-type computation more like multiplication
or division than the usual excitation-inhibition analogous to addition-subtraction).
Kuffler had worked on the third layer of cells in the chain between photoreceptors and brain (he'd
have worked on the photoreceptors and second-order cells, but they were too difficult with 1953
techniques). Hubel and Wiesel discovered a gold mine when they started looking at the brain
structures farther along in the visual pathways, the fourth-order cells in the thalamus, and the
fifth- and higher-order cells in the cerebral cortex. Other neurophysiologists were busy
investigating how sensations from the skin and ears were analyzed in the brain, but the same
techniques applied to the visual pathway had an important advantage: The visual system, more
so than any other of our senses, has been streamlined by one evolutionary adaptation after
another. And we humans tend to think visually, compared to most other animals, giving human
neurophysiologists an advantage in trying to understand such stages of analysis of what the eye
initially "sees" with the first-order photoreceptors.
We understood how cameras worked, and how television cameras worked, and some of us
probably expected the eye to produce a picture of what it saw for presentation to a viewer back in
the brain somewhere. Though we knew that the fine mosaic of about 100 million photoreceptors
receiving the optical image had to repackage some of that information, because there are only a
million "wires" leading back to the brain, we somehow still expected that a faithful image would
be recreated at the other end, much as a television tube recreates what the TV camera saw and
encoded in radio waves.
But there was that troubling result from the frog studies, all that more-detailed-than-a-fly
information that was simply thrown away. Kuffler had fortunately studied the cat's retinal
ganglion cells rather than the frog's (he needed a big eye because of the size and complexity of
the experimental apparatus); clearly, they were at least reporting the boundaries between all
objects. Potentially, the cat's brain ought to be able to read a newspaper. And as Hubel and
Wiesel soon showed, a monkey's retinal ganglion cells were rather like a cat's, except for being
better at fine resolution and at color. What, however, happened in the brain cells to which the
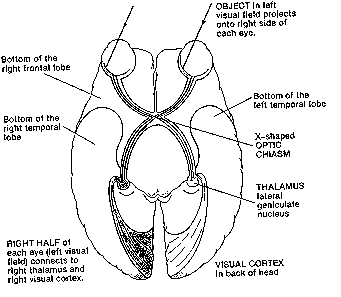
optic nerve reported? The main line from eye to brain goes through the thalamus, whose lateral
geniculate nucleus is an elaborately layered structure (bent knee-like, hence geniculate) that had
long intrigued neuroanatomists. Surely something fancy happened there.
Hubel and Wiesel recorded from those geniculate cells. And at first glimpse and even second
glimpse, they were indistinguishable from the retinal ganglion cells upstream from them --
nothing new seemed to be happening, as if they were just some sort of relay station (this
terminology is borrowed from the old Pony Express, where tired horses were exchanged for
rested ones, the messages in the saddlebags passed on unaltered). The geniculate's receptive
fields were the same center-surround doughnut shapes as the dual-funnel arrangement had
produced in the retinal ganglion cells. About all that the first-pass analysis of the geniculate
showed was that diffuse light was an even poorer stimulus than it was to the retinal cells:
Excitation and inhibition tended to totally cancel out in the geniculate cells, whereas retinal cells
often had some net excitation or inhibition when simply looking at an unbroken field like a
cloudless blue sky. Not until one uses colored lights as stimuli does one discover the geniculate
cells doing something that retina doesn't do: In some cells, the center-surround organization
disappears.
When Hubel and Wiesel looked up in the cerebral cortex at some of the still higher-order cells in
the processing chain, things were obviously quite different than in the retina or geniculate.
Cortex was no relay station; the messages were rearranged here to make new patterns. Most
obviously, the circularly symmetric receptive fields of the cells feeding into the cortex were
somehow modified to make elongated receptive fields. The optimal stimulus for retinal and
geniculate cells was a white spot on a dark background (or in other cells, a black spot on a light
background) of a certain size -- but round, always round. In the cortical cells, round spots might
evoke a response, but the best stimuli were lines and elongated edges.
Now given that switching from spots to lines and edges decreased the responses of the geniculate
cells forming the input to the cortex (compared to the optimal spots), better responses to lines
was a surprise. But a given cortical cell arranged those inputs so as to sum together the activity
of many input cells whose receptive field centers were not all in the same place: Indeed, they
were all strung out in a line, thanks presumably to the genetic instructions that had wired up the
thalamus and cortex during prenatal development.
While some cortical cells obviously preferred horizontal white lines, others preferred vertical
ones. And others preferred various in-between angles, so long as they stayed within about 5° to
10° of their optimal angle. As the tilt of a line shifted (as, for example, the horizon tilts when the
Vineyard ferryboat rolls in a wave), one group of cells fell silent but another group sprang into
activity. Each cell seemed to be a specialist in a particular tilt.
And while some cells were best at edges such as the sea-sky boundary, some preferred narrow
black lines, and others liked narrow white lines (just as one might predict from the two major
classes of their input's Mexican hat arrangements, regular and inverted). So the cortex had a lot
of variety, but it was orderly: Neighboring cells tended to have the same preferences for tilt, until
suddenly reaching a neighbor that liked a quite different tilt. In all of the layers of the cortex, the
tilt preference was the same -- so if one sampled one neuron after another, they would all like the
same tilt. But sometimes the recording probe would stray into an adjacent "column" that
preferred a different tilt -- and so we began to talk about "orientation columns" in the cortex.
Because a cell would respond somewhat to a small spot of light, one could patiently explore its
receptive field, making a "map" of it from which one could predict the optimal stimulus: a
narrow black line at 45°, or an edge oriented vertically, and so forth. And when they tried out
the predicted optimal stimulus, the cell indeed sang in its loudest voice. But some cells refused
to respond to small spots, and so Hubel and Wiesel tried lines and edges -- and they'd usually
work, once they twisted them around to the preferred tilt. It was as if these cells simply had a
high threshold, as if they require a ten-thousand-dollar balance before paying interest. Spots
simply aren't big-league hitters by the time that the information gets to the cortex, according to
this interpretation.
Then they got another surprise: Some such cells would respond to the line even if they moved it
sideways somewhat. For every cell in retina, geniculate, and (so far) cortex that had been
examined, that maneuver would cause the cell to become uninterested, shutting up because the
stimulus had wandered out of its receptive field center or off onto a weak portion of its periphery.
While they couldn't move the tilted line simply anywhere on the retina and have it work, there
was a region perhaps 10-15° wide where the cell still responded to the line. But try tilting the
line away from its favored orientation, and the cell would promptly shut up, no matter where the
line's center was located. Hubel and Wiesel called these "complex cells," and the ones with the
predictable maps "simple cells."
This was an exciting development, as the complex cells appeared to be "generalizing" on the
concept of "line tilted at 45°" regardless of location. Psychologists had long been pushing
generalization as a difference between lower and higher animals: Some species will learn to treat
an erect and an upside-down triangle as "the same thing," and other species will always treat
them as different, refusing to "generalize on the concept of triangle." Complex cells were
generalizing, not about triangles but about one of the parts of the triangle, a tilted line.
So were there higher-order cells in the brain that specialized in triangles, no matter what their
size or tilt, no matter whether black-on-white or white-on-black, no matter whether the center
was filled or not? Were there triangle detectors in higher animals' brains, just as frogs had good
"bug detectors" even out in their retinas?
We know already how to hear isolated cells among the millions of elements in the
system. I cannot imagine a more important task than the reconstruction of the
symphony.
the neuropsychologist Hans-Lukas Teuber
STEPHEN KUFFLER NEVER WON the Nobel Prize, despite his involvement in this string of
successes in uncovering one fundamental aspect after another of how inhibition is used. The
betting among the neurobiological community in the late 1970's was that the next Nobel Prize in
matters neurobiological would go to the triumvirate of Kuffler, Wiesel, and Hubel. But Nobel
Prizes are never awarded posthumously, and Steve Kuffler died while working at his desk in his
Woods Hole home on 11 October 1980, at the age of sixty-seven. As was his custom, he had
been swimming at Stony Beach that morning.
In 1981 Hubel and Wiesel did indeed receive the Nobel Prize in Physiology or Medicine. And as
in all such matters, they were particularly noteworthy representatives of a whole community of
successful scientists. Neurobiology grew up in those years: There was no such word in 1959,
just various neurophysiologists and neuroanatomists and neuropharmacologists and
developmental specialists, etc., coming from backgrounds in medicine, biology, psychology, and
the physical sciences. Today, students can find a variety of graduate programs in neurobiology,
and there are even a few places with undergraduate majors in neurobiology. But mostly we
remain a diverse group of emigrants from other fields, something of a melting pot. Everyone
learns the reductionist techniques, as they remain the best way to train graduate students, but a
few wander off into trying to piece it all together, to see the big picture, maybe discover some
emergent properties.
What we get on the retina, whether we are chickens or human beings, is a welter
of dancing light points stimulating the sensitive rods and cones that fire their
messages into the brain. What we see is a stable world. It takes an effort of the
imagination and a fairly complex apparatus to realize the tremendous gulf that
exists between the two.
Consider any object, such as a book or piece of paper. When we scan it with our
two eyes it projects upon our two retinas a restless, flitting pattern of light of
various wave lengths and intensities.... But we are never conscious of the
objective degree of all these changes unless we use... a peephole that makes us
see a speck of color but masks off its relationships. Those who have used this
magic instrument report the most striking discoveries. A white handkerchief in
the shade may be objectively darker than a lump of coal in the sunshine. We
rarely confuse the one with the other because on the whole the lump of coal will
be the blackest patch in our field of vision, the handkerchief the whitest, and it is
the relative brightness that matters and that we are aware of. The coding process
begins while en route between the retina and our conscious mind.
the art historian E. H. Gombrich, 1959
Email ||
Home Page || Table of Contents || Endnotes for this chapter || continue reading Next Chapter

