|
COPY-AND-PASTE CITATION
copyright ©2000 by William H. Calvin and Derek Bickerton
The nonvirtual book is available from amazon.com or direct from MIT Press. |
William H. Calvin
University of Washington
|
|
I=m not sure if we want, on the analogy to lateral language areas, to call it the Amedial language area,@ however B medial speech area, perhaps, but these vocalizations scarcely meet our criteria for words, let alone structured language. I=m not just being picky about my terminology, however; I suspect that our kind of language didn=t arise from some intensification of the usual ape vocal repertoire.  There are several reasons to suspect this. Besides the distance (almost
half the brain=s
width) between the two areas, there=s
also the meaninglessness of our phonemes, in contrast to the
one-sound-one-meaning obligatory interpretation of chimpanzee vocalizations. I
have trouble imagining how evolution managed the trick of making some new
vocalizations meaningless while retaining the obligatory meaning of the rest,
during the transition period. It happened somehow, of course, but it makes me
think that the meaningless additional ones didn=t
grow up beside the old ones in the competition for cortical space, that they
got a fresh start somewhere else, perhaps not even as vocalizations. I wish we
knew whether the chimps and bonobos had some intermediate specializations,
especially the language-reared individuals. (Do they borrow some lateral
cortex, as they come to understand all of those human sentences? Tune in next
year, I=m
told.) There are several reasons to suspect this. Besides the distance (almost
half the brain=s
width) between the two areas, there=s
also the meaninglessness of our phonemes, in contrast to the
one-sound-one-meaning obligatory interpretation of chimpanzee vocalizations. I
have trouble imagining how evolution managed the trick of making some new
vocalizations meaningless while retaining the obligatory meaning of the rest,
during the transition period. It happened somehow, of course, but it makes me
think that the meaningless additional ones didn=t
grow up beside the old ones in the competition for cortical space, that they
got a fresh start somewhere else, perhaps not even as vocalizations. I wish we
knew whether the chimps and bonobos had some intermediate specializations,
especially the language-reared individuals. (Do they borrow some lateral
cortex, as they come to understand all of those human sentences? Tune in next
year, I=m
told.)
Let me put all of this specialized real estate in a broader context, that of the cerebral cortex in general. It =s a layered structure, mostly up on the surface of the brain abutting the skull, except for what=s buried in infolding. The word Asubcortical@ refers to everything else in the brain. Most of our cortex is newer, mammalian-style neocortex, with six thin layers like a croissant (older parts of cortex, such as hippocampus, have only three, like cakes). And it=s neocortex that has the reputation for handling novelties, such as associative memories. Subcortical structures have more of the reputation of handling subroutines, and with repetition, what is initially a cortical task may become a subcortical one. Tasks like language, full of novelty and multimodality relationships, rely heavily on the neocortex, even though coordinating performance may additionally involve structures like the thalamus and cerebellum. I=m going to stick to neocortical novelty and not attempt to make a wiring diagram of everything involved in aphasia and routine speech. There are lots of ways to describe the real estate; motor strip (that map of the muscles at the rear of the frontal lobe) is an obvious output pathway, and there are obvious sensory input specializations such as the primary somatosensory, primary auditory, and primary visual areas. Everything else (about 85 percent of the brain=s surface area) got called Aassociation cortex.@ It=s not quite terra incognita, however. We started finding a number of secondary sensory maps in parts of association cortex; for vision, there are a few dozen additional maps of the visual world, some quite small.
There =s really no Aexecutive cortex,@ a spot where the big picture comes together and decisions are made. But you won=t go far wrong if you think of your brain as always preparing for action, trying to guess what happens next, and gathering sensory information in aid of tentative plans for action. To the classical who, what, where, when, why, and how, there needs to be added a seventh question: action?This is, you understand, a motor systems neurophysiologist speaking; others often adopt a more inside-looking-out psychological perspective. The difference in perspectives often comes to the surface when discussing consciousness; while I =m busy discussing how Darwinian processes sort through ambiguity and create novel plans of action, they usually are talking instead about awareness, how you focus your attention on various aspects of the environment.As Gregory Bateson once said, information is data that makes a difference B that which sways your choice about what to do next. Sensory information is sometimes collected systematically or at random (we are, to some extent, Ainterrupt driven@). But much of the time, information from the sense organs and our memories is being collected in aid of a plan of action, one of several that are percolating up into consciousness. Each possible action triggers a number of questions, including the aforementioned who-what-where-when-why-how.And that preparation for action perspective helps us to understand verbs and their needs for linking to themes Bagents, beneficiaries, and so forth. As I mentioned earlier, verbs seem to live in the frontal lobe behind the temple. The most elementary verbs naturally come with arguments. A verb like Ago@ always needs an orientation: when go surfaces as an issue, secondary issues need to be decided involving where to go, toward what object or person, around what obstacles, with what means, and so forth. A verb like Agive@ immediately raises such issues as Agive what@ and Ato whom,@ even if the giver is yourself. Because the frontal lobe is also engaged in marshaling actions over some time span (long holds, for example, are a prefrontal specialty in monkeys doing a delayed-match-to-sample task), such ancillary issues as when to go may be closely linked. That=s why I said adverbs such as Aquickly@ and Aslowly@ likely resided in frontal lobe as well, though there=s not much data yet.And, of course, we =re really talking about connotations of a word here, not its pronunciation. Sometimes a stroke patient can use a word like crack as a verb (Acrack a nut@ if they have an intact frontal lobe) but not as a noun (Aa crack in a window@ if they have temporal lobe damage).Nouns and adjectives involve a different type of concept; they may make links but usually not obligatory ones in the demanding manner of verbs. There has been a major rearrangement of the temporal lobe that features space for concepts, one that may have something to do with the vocabulary-without-structuring stage of protolanguage. Some stroke patients can name tools, but not animals. Others are missing their plants, or body parts, or verbs, B or perhaps the combination of food, fruits, and vegetables. Mind you, they can usually recognize the word, and write the word. What they are usually missing is the visual representation of that category.The secondary visual areas in the parietal and temporal lobes feature a where-what split, with the parietal lobe being more concerned with where, and the temporal lobe with the what issues. In monkeys, much of the temporal lobe is devoted to the what-type secondary visual areas; the top side of the temporal lobe is auditory. (I =m going to have to compare monkeys to humans because no one yet knows about the ape temporal lobe, though they=re starting to train one of the language-reared chimpanzees in a context that allows brain scans.)
So far as neurologists can tell from studying stroke patients, the reorganized human temporal lobe is all about categories, exactly what Derek needs for protolanguage =s vocabulary. Toward the rear are such things as color, then comes stuff more like tool concepts. The front end of the temporal lobe (Atemporal tip@ seems to have more to do with carefully delimited categories of one: proper nouns. Epileptic patients who have had a temporal tip surgically removed sometimes say that they=re always having to write down a new name, that their recall of proper names isn=t what it used to be, pre-op.There =s another way of studying concepts: try to confuse a small locality in the cortex by tingling it with a mild electrical current. The opportunities to get such data are largely come from the surgical treatment of epileptic seizures and the associated pre-op workups needed to make sure that the patient will benefit from surgery. I=ll save you the details (they=re in Conversations with Neil=s Brain that George Ojemann and I wrote together) and just say that the brief stimulation doesn=t evoke concepts or words, it merely prevents them from arising, despite the patient=s attempt to access them. The sensitive spot for disrupting a tool concept might be no larger than a small coin (strokes usually have to be larger than that just to be noticed at all; stimulation can identify much smaller specialized regions).There are other cortical locales known as Anaming sites@ because stimulation at them causes the patient to fumble at naming any sort of common object: shown a drawing, the patient says, AThis is a . . . a . . . . Oh, I know what that is, it=s a . . . .@ The patient can speak, but can=t get out the common name; turn off the current, and the name will often pop out, finally. A half-dozen such naming sites can be found in a given patient, scattered around the temporal, parietal, and frontal lobes on the lateral surface; they=re different places in different people, but usually not more than several fingers= width from the left sylvian fissure (there are probably more, buried in folds, inaccessible). Neurosurgeons are very careful to avoid these naming sites when removing epileptic tissue from the brain, for fear of creating an aphasia (a loss of language abilities). If there has to be a tradeoff, aphasia is usually considered worse than epilepsy.One fascinating aspect of naming sites is that, in bilingual patients, they are sometimes different for the different languages. Test the patient with a series of object drawings, asking him to name them in English, and find the coin-sized sites where stimulation blocks the naming ( Anaming sites@). Then repeat the series, asking him to name them in Spanish, or whatever. Some of the English naming sites will not block naming in Spanish; there will also be some new naming sites, where naming in Spanish is blocked but not in English. Some naming sites block both languages. The naming sites are only a small percentage of all the sites tested (imaging methods show that much wider areas of cortex are working harder during such tasks B they=re involved but not essential). The momentary electrical buzz can be used to study a number of different
brain functions, a technique pioneered in middle of the twentieth century by
the Montreal neurosurgeon Wilder Penfield, and greatly expanded over the last
three decades by George Ojemann in Seattle. In most cases, it causes a
function to fail, though in the motor cortex in the back end of the frontal
lobe, it causes poorly coordinated movements. (AHey,
somebody moved my right thumb!@)
Crude sensory impressions can be evoked in primary sensory cortices, but
usually not formed sensations. The reports of hearing music, or seeing a man
walking a dog, may be related to the patient=s
typical aura of an impending seizure B
something that the electrical buzz can provoke, if too strong. So these
so-called Aexperiential
responses@
are not necessarily normal ones, but perhaps those Aburned
in@
by previous seizures B
set pieces, not easily disassociated into meaningful parts, rather like
multi-word exclamations or whale songs. The momentary electrical buzz can be used to study a number of different
brain functions, a technique pioneered in middle of the twentieth century by
the Montreal neurosurgeon Wilder Penfield, and greatly expanded over the last
three decades by George Ojemann in Seattle. In most cases, it causes a
function to fail, though in the motor cortex in the back end of the frontal
lobe, it causes poorly coordinated movements. (AHey,
somebody moved my right thumb!@)
Crude sensory impressions can be evoked in primary sensory cortices, but
usually not formed sensations. The reports of hearing music, or seeing a man
walking a dog, may be related to the patient=s
typical aura of an impending seizure B
something that the electrical buzz can provoke, if too strong. So these
so-called Aexperiential
responses@
are not necessarily normal ones, but perhaps those Aburned
in@
by previous seizures B
set pieces, not easily disassociated into meaningful parts, rather like
multi-word exclamations or whale songs.
Where, Derek will surely ask, are the boundary words located? Words like Asince@ and Abecause,@ that signal a new verb coming up? Or words like Aof@ and Ainto@ that signal a new phrase? There are only a few dozen such words, a closed class that is difficult to expand (though we can always add more nouns and verbs). If the brain handled a closed-class word as a special case (in the manner of irregular verbs and nouns) in a particular place, we would have a substantial clue to how a structured sentence is parsed into smaller units.Alas, the closed-class words have not usually been part of the experimental design for cortical localization of language, at least not thus far. Ojemann and coworkers do see cortical Agrammar@ sites where the patient makes mistakes when reading aloud, substituting the wrong verb endings, pronouns, conjunctions, and prepositions, but at these sites, patients don=t make mistakes reading nouns and verb stems.The loss of cortical specializations for the closed class might be important for Broca =s aphasia, where language production tends to degenerate into protolanguage=s short utterances. Such patients often have trouble with sentence understanding, typically with the closed-class words such as conjunctions and prepositions, even though their understanding of other word types remains.In the Wernicke-style aphasias, understanding is often poor even though production retains phrases and clauses. Such patients talk fluently, even excessively, but they sometimes use words that make no sense. So quality isn =t very good, in the manner that might be expected of a Darwinian competition that was never able to operate for enough generations before the utterance was put into action or comprehension deemed good enough.DB: This is very striking evidence for what I think you are going to say later B that there is Darwinian competition between sentences, a process that surprisingly produces Agarbage in, quality out.@ Because if there weren=t, where would these nonsense phrases come from? What they often look like are bits of quite different sentences put together, like this one: AI=ll tell I=ve been my wife was every time dollars for teeth and my wife didn=t pay any at home.@ Aphasias come in many varieties. What students, fresh from learning about sensory areas and movement areas elsewhere, tend to remember about them is a great oversimplification, but here it is: understanding depends a lot on the cortical areas bordering the rear half of the left sylvian fissure, while expression is more a matter of frontal lobe areas somewhat in front of the temporal tip. I shudder to think of all of the exceptions to this Arule.@During the first century of neurology, the aphasias provided an insight into cerebral localization of function for language via their correlation with surgical and postmortem findings. But the aphasia subcategories which were invented then have not, so far, meshed well with those from the more modern physiological techniques. One reason is that the cortical stimulation technique revealed that the areas surrounding the sylvian fissure specialize in both sensation and movement, at least when novel sequences are involved. The experimental design for stimulation mapping tried, for this case, to avoid issues of language understanding by using sound sequences and movement sequences that were linguistically meaningless. For example, the patient sees a cartoon sequence of three frames, with one of the experimenters modeling three oral-facial postures: puff out cheeks, stick out tongue, clench the teeth. And so the well-trained patient does all three actions, in that order. Another cartoon slide comes up, and the patient does another three. Stimulation occurs during some slides, but not during others; it=s so weak that he doesn=t know when it is occurring. During stimulation, he may produce the movements, but in the wrong order. Or he may add movements not modeled on the current slide. If the cartoon series is simply the same posture repeated three times, the patient makes no errors. Stimulation is only disrupting when a sequence of different movements must be produced.
In a rim around the perisylvian sequencing core, there are a number of sites in the frontal, parietal and temporal lobes where stimulation disrupts short-term memory tasks. But first I =d better explain some of the current concepts of how memory works in the brain, because it isn=t much like the way it works in a computer. |
 These days, we tend to talk about the cortex
These days, we tend to talk about the cortex Overall, of course, the human brain is a lot bigger than monkey brains.
Monkeys have about enough cerebral cortex on both sides to cover a postcard,
were the thin cortical rim peeled off and flattened out. Apes (that
Overall, of course, the human brain is a lot bigger than monkey brains.
Monkeys have about enough cerebral cortex on both sides to cover a postcard,
were the thin cortical rim peeled off and flattened out. Apes (that But the human cortex is rearranged, too, and temporal lobe is the
best-studied case. The human temporal lobe has all of those monkey-style
visual areas back in the rear, the primary auditory area on the top middle,
with something else going on in the middle and front of the temporal lobe.
What filled in?
But the human cortex is rearranged, too, and temporal lobe is the
best-studied case. The human temporal lobe has all of those monkey-style
visual areas back in the rear, the primary auditory area on the top middle,
with something else going on in the middle and front of the temporal lobe.
What filled in?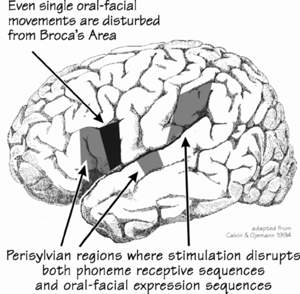 Movements are supposed to be a frontal lobe function, but movement
sequencing seems to involve the temporal and the parietal lobe as well, at
some sites near the sylvian fissure. At about 86 percent of these
movement-sequencing sites, a listening-to-sound-
Movements are supposed to be a frontal lobe function, but movement
sequencing seems to involve the temporal and the parietal lobe as well, at
some sites near the sylvian fissure. At about 86 percent of these
movement-sequencing sites, a listening-to-sound-
 On to the NEXT CHAPTER On to the NEXT CHAPTER
Notes and References for this chapter Copyright ©2000 by William H. Calvin and Derek Bickerton The nonvirtual book is available from amazon.com or direct from MIT Press.
Book's Table of Contents
|