Competere
per la coscienza:
un meccanismo darwiniano a un
livello
appropriato di spiegazione
William H. Calvin
Universitŕ
di Washington
Facoltŕ di Psichiatria e Scienze
Comportamentali
Seattle WA 98195-1800 USA
Abstract.
Trattare la coscienza in quanto consapevolezza o attenzione significa
sottovalutarla enormemente, ignorando i livelli temporanei di organizzazione
associati alle funzioni intellettive superiori, quali sintassi, invenzione,
logica e musica. I compiti che richiedono l’intervento della coscienza
tendono a essere quelli che implicano molte risorse; le attivitŕ di routine
possono essere svolte automaticamente, ma gestire l’ambiguitŕ, cercare
soluzioni “offline”, produrre scelte creative ed eseguire movimenti di
precisione possono richiedere l’impiego di settori di neocorteccia
sostanziali. Su cercherŕ qui di chiarire i livelli di spiegazione appropriati,
dagli aspetti quantici alla dinamica della corteccia associativa, per proporre
un meccanismo specifico (la coscienza come vincitore attuale dei meccanismi di
competizione darwiniana nei processi di copiatura all’interno della
corteccia cerebrale) che sia in grado di includere sia le funzioni superiori
della coscienza che alcuni aspetti attenzionali. Tale meccanismo comprende
anche aspetti quali lo spazio di codificazione appropriato ad analogie e un
processo darwiniano di supervisione che influenzi l’azione di altri processi
darwiniani.
Francis Crick nota che,
se una volta ci si preoccupava del confine tra materia vivente e non, tale
confine sembra ormai aver perso significato, mentre si parla dei diversi
aspetti della biologia molecolare. Oggi i neurologi e i neurofisiologi credono
che l’attuale interesse del mondo scientifico e filosofico per la coscienza
sarŕ presto altrettanto obsoleto, e che si parlerŕ semplicemente dei vari
processi fisiologici deputati all’attenzione e alla soluzione creativa di
problemi. Ciň non significa che il termine coscienza scomparirŕ dal nostro
vocabolario, allo stesso modo in cui la prudenza dei filosofi nei confronti
degli errori dei classici continuerŕ a dimostrarsi utile. Come la parola “vivente”,
anche “conscio” e “mente” resteranno utili scorciatoie; anche quanti
di noi comprendano pienamente tutti i processi fisiologici coinvolti avranno
ancora bisogno della parola “coscienza”1, non solo per
comunicare con i profani del campo ma anche per riuscire a concepire la
materia stessa.
Mi si consenta allora un’introduzione all’argomento da un punto di
vista neurofisiologico, che spero possa essere utile nell’acquisizione di
una maggiore consapevolezza riguardo a quello che Dan Dennet ha definito
“l’ultimo mistero”. Secondo Dennet un mistero č un “fenomeno che non
si č ancora in grado di pensare”. Cercherň qui di chiarire i livelli di
spiegazione appropriati, per proporre successivamente un meccanismo candidato,
una “macchina darwininana” (Calvin 1987, 1996) che sia in grado di
includere sia le funzioni superiori della coscienza che alcuni aspetti
attenzionali; parte di questo articolo č un adattamento da due miei libri,
pubblicati nel 1996: How Brains Think (Basic Books, New York) e The Cerebral Code (MIT Press, Cambridge MA).
La coscienza č la punta dell’iceberg, nel senso che molti
altri processi si determinano nel cervello contemporaneamente, senza che ne
siamo consapevoli. In effetti non siamo in grado di dar conto della maggior
parte delle funzioni svolte dal nostro cervello. Non siamo in grado di
riportare la pressione arteriosa senza una strumentazione esterna, nonostante
sia il cervello a misurare e regolare la pressione sistolica e diastolica;
semplicemente tale misurazione non č esprimibile verbalmente.
Nello
studio del cervello si devono misurare livelli multipli di meccanismi, dalle
sinapsi alle cellule, ai circuiti nervosi, ai moduli, e ancora oltre. Devono
essere misurati anche i livelli multipli di spiegazione fenomenologia, quali
attributi, oggetti, categorie, analogie e metafore. Come gli intermediari sono
una realtŕ comune al mondo economico, cosě ci si aspetta di osservare strati
multipli di rappresentazione tra la nostra coscienza e la realtŕ. Derek
Bickerton ha osservato:
Piů la coscienza č profonda, piů strati di elaborazione
ci separano dal mondo… L’allontanamento progressivo dal mondo esterno č
semplicemente il prezzo da pagare per la conoscenza del mondo stesso. Piů la
coscienza del mondo č profonda e vasta, piů complessi divengono gli strati
di elaborazione necessari per raggiungere tale coscienza.
Esistono comunque treni di pensiero subconsci, ed essi competono per
ottenere “attenzione”; lo strato immediatamente sottostante la coscienza
puň implicare un meccanismo di
competizione.
Sebbene sia spontanea l’analogia con lo
spettatore televisivo che fa “zapping”, e i sogni notturni sembrano spesso
uno zapping continuo da una soap opera all’altra, non č necessario avere un
canale in particolare da cui visionare le scelte. Il canale “migliore”
necessita di vincere solo temporaneamente sugli altri, nella lotta per
l’accesso ai mezzi espressivi, quali il linguaggio e i movimenti del corpo.
Presto un altro canale diventa dominante e si parla di “spostamento
dell’attenzione”, ma un agente che decida o agisca non č indispensabile,
non piů di quanto ne abbia bisogno il ghiaccio per sciogliersi.
L’espressione
non implica movimenti evidenti: il silenzio č uno stato importante, in cui
ragioniamo, creiamo alternative e scegliamo tra esse. La rotazione mentale di
oggetti complessi illustra una possibilitŕ di non linguaggio del pensiero
cosciente, ma da un punto di vista neurofisiologico non c’č niente, in tale
visione d’insieme, che richieda un posto centrale. Il “centro della
coscienza” potrebbe invece sportarsi continuamente dalle aree del cervello
deputate al linguaggio a quelle del non linguaggio, dal lobo frontale a quello
parietale, dall’emisfero sinistro a quello destro, e forse anche dalle
strutture corticali a quelle subcorticali: dovunque sia possibile creare
movimenti originali. Sembrerebbe
quindi possibile svolgere diverse funzioni contemporaneamente, proprio come si
riesce a cucire o ascoltare la radio, mentre si guarda la televisione. Non
tutte le nostre attivitŕ sono controllate dal centro di coscienza attivo in
quel momento. Karl Popper ha osservato che ponderare alternative č
un’attivitŕ particolarmente “cosciente”:
Gran
parte del nostro comportamento intenzionale (e presumibilmente anche degli
animali) si determina senza l’intervento della coscienza… I problemi che
possono essere risolti con comportamenti di routine non richiedono
l’intervento della coscienza; i successi biologici che ne implicano
l’intervento sono soluzioni a problemi
non ordinari. Il ruolo della coscienza risulta piů chiaramente laddove un
obiettivo… sia ottenuto per vie
alternative, e quando si sperimentino due o piů possibilitŕ, in seguito
ad attenta valutazione.
Sebbene pensi che Popper abbia avuto una buona intuizione, io
generalizzerei dicendo che le funzioni che richiedono l’intervento della
coscienza tendono a essere quelle piů impegnative. I compiti di routine
possono essere svolti automaticamente, ma gestire l’ambiguitŕ, cercare
soluzioni, produrre scelte creative ed eseguire movimenti di precisione
possono richiedere temporaneamente l’impiego di settori sostanziali di
neocorteccia.
Coscienza
e livelli di spiegazione
Ci si domanda allora che cosa costituisca una spiegazione. Per
illustare seriamente il significato di coscienza bisogna evitare concetti
matematici che ottengono il risultato di confondere piů che chiarire,
evitando “prove per mancanza di immaginazione”, come quando si stabilisce,
per presunzione e impazienza, che non esistono alternative alle risposte
ottenute. Quando si parla di cervello č essenziale assicurarsi che le nostre
teorie siano impostate al giusto livello di spiegazione meccanica.
L’attivitŕ cerebrale implica almeno una dozzina di livelli, e
trovare livelli appropriati agli aspetti della coscienza non č stato semplice.
Si cerca sempre di estrapolare la propria materia di studio e dichiararne la
pertinenza alla coscienza. Poiché mi accingo a fare lo stesso, mi si consenta
qualche parola sui livelli, in particolare perché essi sono in relazione con
le proprietŕ emergenti e quei “cambiamenti di stato”, per cui č bene sottolineare due punti fondamentali in questo
articolo, che č un breve sunto di due mie pubblicazioni maggiori, How
Brains Think e Cerebral Code, qui
spesso citate. Il primo č una proposta di un meccanismo di
coscienza specifico (come vincitore attuale dei meccanismi di competizione
darwiniana nei processi di copiatura, all’interno della corteccia
associativa), mentre il secondo rappresenta una spiegazione su cosa sia un
livello corretto, a cui cercare un meccanismo della coscienza. Spero che
risulteranno entrambi appropriati, ma il livello di analisi si propone
un’utilitŕ piů ampia: č infatti inteso come critica su molto di quanto
sia stato scritto sull’argomento.
Ciň che manca in gran parte dei dibattiti sulla coscienza č,
sorprendentemente, l’idea stessa che esistano livelli di spiegazione.
Douglas Hofstadter (1985) offre un buon esempio di livelli nell’osservare
che la causa di un ingorgo stradale non dipende da una sola auto o dai suoi
componenti; gli ingorghi stradali sono un esempio di auto organizzazione, piů
facilmente riconoscibile quando il meccanismo di partenza e di stop raggiunge
un livello estremo di semi-stabilitŕ,
quella cristallizzazione nota come ingorgo. Un ingorgo stradale occasionale puň
essere dovuto al malfunzionamento dei componenti, ma le candele di accensione
non sono un illuminante livello di analisi, non se paragonato
all’amalgamarsi del traffico, ad ampi parcheggi, ai tempi di reazione
dell’automobilista, alla posizione della segnaletica e alla mancata
accelerazione da parte dell’automobilista a causa di una collina o un ponte
rialzato. I livelli di spiegazione piů elementari sono in gran parte
irrilevanti nella formazione di un ingorgo stradale. Tale disaccoppiamento
(decoupling) fu enfatizzato dal fisico Heinz Pagels (1988), il quale osservň:
“Il
disaccoppiamento causale” tra i livelli del mondo implica che per capire
le basi materiali di certe regole si deve procedere al livello immediatamente
inferiore; ma tali regole possono essere applicate tranquillamente senza
alcuna referenza ai livelli piů elementari. E’ interessante notare come la
divisione delle scienze naturali rifletta tale disaccoppiamento causale. La
fisica nucleare, la fisica atomica, la chimica, la biologia molecolare, la
biochimica e la genetica sono ciascuna discipline indipendenti, valide per se
stesse, conseguenza del disaccoppiamento causale tra di esse… Tale serie di
“disaccoppiamenti causali” puň risultare straordinariamente complessa e
intricata, oltre quanto si possa attualmente immaginare. Eppure alla fine sarŕ
possibile formulare una teoria della mente e della coscienza, in cui la mente
č talmente disaccoppiata dai propri sistemi di supporto materiale che sembra
esserne indipendente, per poi “dimenticare”
come ci si č arrivati… Il fenomeno biologico della coscienza
autoriflessiva č semplicemente l’ultimo di una lunga e complessa serie di
“disaccoppiamenti causali” dal mondo della materia.
Stabilitŕ stratificata ed emergenti
Strettamente correlata č
la nozione di proprietŕ emergenti: ingorghi stradali e cristalli emergono da
combinazioni e ci si aspetta che l’emergenza abbia un ruolo importante nei
livelli di organizzazione transitori coinvolti nelle funzioni intellettive
superiori (linguaggio, inventiva, giochi, ecc.). Nella ricerca di un livello
corrispondente alla coscienza, č bene ricordare che i livelli derivano da
quella che Jacob Bronowski (1973) ha definito stabilitŕ stratificata:
La
natura procede per gradi: gli atomi formano molecole, le molecole creano basi,
le basi dirigono la formazione di aminoacidi, gli aminoacidi formano proteine
e le proteine lavorano in cellule. Le cellule presiedono alla formazione prima
degli organismi piů semplici e poi di quelli piů complessi, salendo un
gradino alla volta. Le unitŕ stabili che compongono un livello o uno strato
sono la materia prima per incontri casuali che producono configurazioni
superiori, alcune delle quali potrebbero essere stabili…
L’evoluzione consiste proprio nel salire questa scala dai gradini piů
semplici a quelli piů complessi, ciascuno dei quali ha una propria stabilitŕ.
Il tumulto di combinazioni casuali produce occasionalmente un nuovo tipo
di organizzazione: alcune forme, ad esempio le cellette esagonali che si
formano nella polenta se si dimentica di amalgamarla, sono effimere, come i
contenuti della coscienza. Altre forme possono avere una sorta di “dente
d’arresto” che impedisce una regressione una volta raggiunto il nuovo
ordine. Se i cristalli sono le forme semi-stabili meglio conosciute, le
strutture molecolari ne rappresentano un’altra varietŕ, ed č anche
possibile che ci siano forme semi-stabili ai livelli intermedi, come gli stati
quantici di formazioni microtubulari, dove i fisici della coscienza vorrebbero
si svolgesse l’azione della coscienza.
“Di
conseguenza, il livello neuronale di descrizione che fornisce l’attuale
immagine del cervello e della mente č solo un’ombra
del piů profondo livello di azione citoscheletrica, ed č a tale livello che
va cercata la base fisica della mente!”
Roger
Penrose, Shadows of the Mind, 1994
Seppure io speri che gli effetti quantici sulla
coscienza siano reali (come si noterŕ quando passerň alle possibilitŕ
tecnologiche), tale ipotesi implica perň una serie di problemi, tra cui di
non minore importanza č la scarsa evidenza che esista una connessione tra i
campi quantici e il sincronismo corticale, tra il sincronismo e i vincoli di
percezione, tra i vincoli e l’attenzione, o tra l’attenzione e la molto piů
vasta serie di fenomeni collegati alla coscienza; tale parola ha molte
connotazioni e sicuramente non tutte sono coinvolte nello stesso meccanismo.
Per
quanto riguarda il primo passaggio dai Qm (la mia abbreviazione per tutti i
“quanti” esaminati da Jibu e
Yasue, 1995) al sincronismo, si dovrebbe sempre essere prudenti verso ogni
meccanismo che vanti di attraversare una dozzina di livelli di organizzazione
per produrre un effetto straordinario. Come ho spiegato in How Brains Think:
[Un]
livello di indagine sulla coscienza piů appropriato č probabilmente a un
livello di organizzazione immediatamente sottostante a quello della percezione
e dell’inventiva: secondo la mia ipotesi č probabile che i circuiti nervosi
della corteccia cerebrale e un’auto organizzazione dinamica implichino
modelli di accensione in una serie di regioni corticali delle dimensioni di un
francobollo e in costante cambiamento. La coscienza non si trova certo nelle
“cantine” della chimica o della fisica, in nessuna accezione. Il tentativo
di passare, in un unico salto, dal seminterrato della meccanica dei quanti
all’attico della coscienza č ciň che io chiamo “il sogno del Portinaio”.
La meccanica dei quanti č probabilmente essenziale alla coscienza nello
stesso modo in cui i cristalli una volta erano essenziali alle radio, o le
candele di accensione sono tuttora essenziali agli ingorghi stradali.
Necessari ma non sufficienti. L’argomento č interessante in sé, ma č
soltanto lontanamente legato alla vita mentale, e non va confuso con i livelli
temporanei di organizzazione associati alle funzioni superiori.
Bootstrapping
darwiniano della qualitŕ
Una
prova di esistenza per una buona alternativa č potenzialmente molto piů
persuasiva della migliore critica sull’essenza della meccanica dei quanti o
della vocina interiore, quindi mi si lasci cambiare marcia a questo punto. Se
decidiamo che la coscienza č importante per ciň che Popper ha definito “la
soluzione ai problemi che non siano di routine”, allora creare un corso di
azioni “dal buono all’eccellente” (“qualitŕ”) a livello di pensiero
rappresenta un’aspetto chiave della coscienza, che va ben oltre la mera
consapevolezza o gli spostamenti di attenzione.
Č ormai sicuro che esistano meccanismi capaci
di effettuare un bootstrapping ricorsivo di novitŕ casuali verso qualcosa di
qualitŕ. Negli ultimi 160 anni, si č avuta una prova di esistenza, il
processo darwiniano (Calvin 1996, 1997). Il modo in cui si raggiunga la qualitŕ
č stato a lungo un problema per le menti migliori della biologia evolutiva
(Maynard Smith e Szathmáry 1995), e la lenta evoluzione della specie, su una
scala temporale di millenni, non ne č piů l’unico esempio: la risposta
immunitaria č un altro processo darwiniano, che opera su una scala di giorni
e settimane, mentre anticorpi progressivamente migliori si formano in risposta
alla sfida di un nuovo antigene. Per decenni le scienze informatiche hanno
usato un procedimento di ricerca di soluzione chiamata algoritmo genetico
(Holland 1992), che imita una versiona allargata del processo darwiniano su
una scala temporale limitata solo dalle dimensioni e dalla velocitŕ dei
computer.
Sarebbe
sorprendente se il cervello non facesse uso di tale fondamentale principio di
bootstrapping della qualitŕ. Ci si domanda allora se questo ben noto processo
possa operare nel cervello su una scala temporale pari a quella del pensiero e
dell’azione e se possa spiegare gran parte di ciň che chiamiamo “coscienza”.
Mi sono impegnato a rispondere a tali quesiti in uno dei libri che ho
pubblicato nel 1996, The Cerebral Code, che analizza i circuiti di
neuroni eccitatori ricorrenti della neocorteccia dei mammiferi. Ho dimostrato
che il diffuso principio del sistema di comunicazione elettrico era capace di
passare attraverso i sei elementi
essenziali del processo darwiniano classico:
un modello
(in questo caso il modello di accensione spazio-temporale di un
assemblaggio cellulare Hebbiano) che copia
con una variazione occasionale, dove le popolazioni delle varianti competono
per un ristretto spazio di elaborazione, e il cui relativo successo č
influenzato da un ambiente ricco di
sfaccettature, sia memorizzato che in tempo reale, in questo caso, e con
ulteriori variazioni che convergono sull’esemplare di
maggiore successo della generazione attuale (il principio di ereditarietŕ
di Darwin).
Tale processo darwiniano ormai collaudato č ciň che si associa allo
sviluppo ricorsivo della qualitŕ, che non andrebbe confuso con la mera
selezione di un singolo modello e altri “insiemi sparsi”, che utilizzano
solo alcuni dei sei “elementi essenziali” (Calvin 1997).
Il modo migliore per dimostrare ciň che io
considero un livello di spiegazione appropriato, ossia quello dei circuiti
nervosi che coinvolgono settori di corteccia cerebrale delle dimensioni di un
francobollo (abbiamo abbastanza corteccia da riempire quattro fogli A4), sta
nel portare al lettore un esempio concreto. Non si tratta dell’unico esempio
possibile, semplicemente č quello che si deve a Darwin, ma mi aspetto che
anche altre teorie candidate arriveranno a utilizzare lo stesso livello di
spiegazione, quello delle proprietŕ emergenti nella dinamica dello
spostamento di settori corticali sostanziali. Dopo aver spiegato i circuiti
nervosi in misura tale che il lettore possa immaginare il funzionamento di un
processo darwiniano nella neocorteccia, tornerň a parlare brevemente della
Coscienza Quantica, dei treni di pensiero e di una copertura esplicativa
indispensabile a una teoria della mente.
Circuiti
nervosi darwiniani
I circuiti nervosi
corticali che rendono un collaudato processo Darwiniano possibile non sono un
elemento oscuro, noto solo a pochi neuroanatomisti: possono essere considerati
facilmente il piů importante principio di comunicazione elettrica osservabile
nella corteccia cerebrale, quello delle connessioni eccitatorie ricorrenti tra
neuroni piramidali limitrofi, negli strati superiori della neocorteccia. Č
occorso un po’ di tempo per capire una delle implicazioni che ne derivano,
ossia una proprietŕ emergente del sistema di circuiti nervosi che nessuno dei
singoli elementi possiede: occasionalmente, ci si aspetta di osservare una
serie di insiemi triangolari sincronizzati di neuroni piramidali, con nodi a
distanza di 0,5 mm.
Ciascun neurone piramidale
ha un assone che si dirama 10.000 volte; alcune di tali diramazioni viaggiano
nella sostanza bianca, ma la maggior parte non lascia mai gli strati corticali,
terminando in una sinapsi di un millimetro circa. L’assone viaggia lateralmente, eccitando gli altri neuroni
corticali, per lo piů altri neuroni piramidali. I neuroni piramidali degli
strati profondi (V e VI) hanno assoni laterali che rimangono negli strati
corticali, alcuni terminano nelle vicinanze
e altri piů lontano.
Sono comunque i contatti elettrici ossevabili (cfr.
Lund e altri autori, 1993) nei prolungamenti assonici dei neuroni piramidali
dello strato superficiale (strati II e III) a essere sorprendenti: i terminali
sono modellati, gli assoni sono paragonabili a treni espresso che non si
fermano a una lunga serie di fermate intermedie, concentrando la propria
azione sinaptica in zone distanti circa 0,5 mm, e rendendo cosě possibile la
formazione occasionale di insiemi triangolari sncronizzati.
Nonostante lo spazio di 0,5 mm sia simile a
quello delle macrocolonne (ad esempio, le colonne di orientamento della
corteccia visiva) e nonostante questo possa contribuire all’organizzazione
di tali macrocolonne durante lo sviluppo (Calvin, 1995), le analogie con le
macrocolonne, cosě come le conosciamo oggi, sono limitate. L’aspetto
intrigante č la loro relazione con le minicolonne (le colonne di orientamento
sono l’esempio meglio conosciuto), le quali sono colonne verticali composte
da circa un centinaio di neuroni, organizzate attorno a un fascio dendritico,
come gambi di sedano; la distanza tra fasci limitrofi č di circa
0,023-0,031mm nella scimmia, il doppio nel gatto (Peters and Yilmaz, 1993). Le
azioni assoniche piů vicine di un neurone piramidale superficiale sono a
distanza di 0,5 mm, ma solo rispetto ai neuroni adiacenti. Le cellule di una
minicolonna tendono ad avere “interessi” simili, come negli stimoli visivi
prolungati di un particolare orientamento, e una delle ragioni sta in questa
eccitazione locale ricorrente.
“ANELLO” DI
ECCITAZIONE PREVISTO
|
|
La figura rappresenta parte del neurone piramidale della scimmia,
ricostruito da McGuire e altri (J.Comp. Neurol. 1991), e mostra
i terminali massonici rispetto ai neuroni limitrofi (assoni
sottili nell’albero dendritico) e le ramificazioni verso
cellule distanti una macrocolonna.
La
tendenza degli assoni a “saltare le fermate” come un treno
espresso dovrebbe risultare in un centro di luce intermittente
circondato da un anello di eccitazione.
Poiché
gli altri neuroni piramidali superficiali si comportano allo
stesso modo, grupi di neuroni in un insieme triangolare
tenderanno a eccitarsi a vicenda. Il sincronismo ne dovrebbe
essere una conseguenza occasionale.
|
|
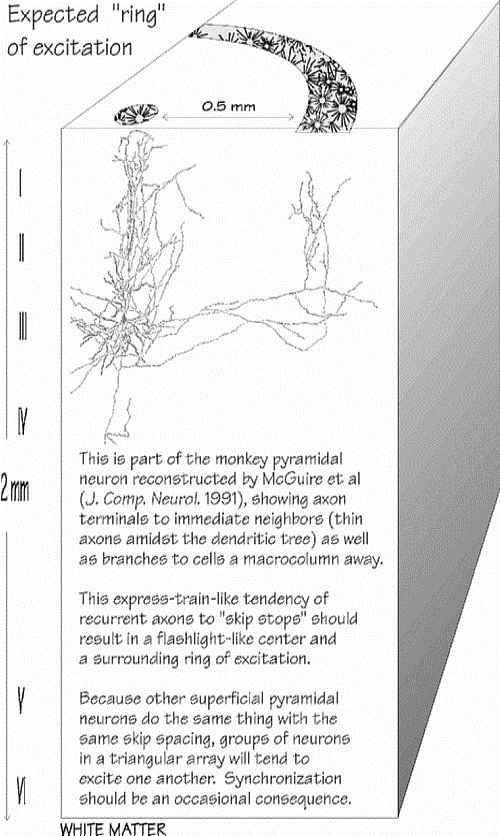
I prolungamenti
assonici piů lunghi tendono a
saltare sinapsi per 0,4-0,6 mm, ma l’analogia col treno espresso č troppo
“monodimensionale” per dare un’idea della realtŕ: i prolungamenti
assonici vanno in molte direzioni radiali. Dall’alto, si dovrebbe osservare
una sorta di anello; mentre i prolungamenti assonici di una cellula non
andranno a riempire l’anello, in una data minicolonna ci sono 39 neuroni
piramidali superficiali che svolgono le stesse funzioni. Si paragoni
l’azione svolta a un fascio di luce mal orientata e intermittente: un
puntino centrale e un anello luminoso nella periferia. Poiché molti assoni
proseguono la corsa per altri 0,5 mm, si devono immaginare anelli aggiuntivi a
intervalli di 0,5 mm, simili a un barilotto, con gradazioni di grigio che
sbiadiscono progressivamente.
Sincronismo
su piccola scala
Conseguenza dell’assone/treno-espresso
č che le cellule a distanza di 0,5 mm tenderanno a comunicare tra loro: si
ecciteranno in maniera ricorrente. In caso di forze sinaptiche elevate, č
possibile che vi sia una sorta di “cappio” che imprigioni i terminali45,
ma anche le forze sinaptiche deboli determinano un importante conseguenza: le
forze di trascinamento. Da quando il fisico olandese Christiaan Huygens notň
nel 1665 che orologi pendolo posti sullo stesso ripiano sincronizzavano il
proprio ticchettio nell’arco di una mezz’ora, si sono compiuti enormi
passi avanti riguardo al trascinamento. In un numero di Science del
1935, Hugh Smith ne riportava un esempio sensazionale dalle Filippine:
Si
immagini un albero alto 10, 12 metri, che abbia una lucciola su ogni foglia,
che tutte le lucciole lampeggino all’unisono circa tre volte ogni due
secondi, e che l’albero si trovi nel buio piů completo, tra un
lampeggiamento e l’altro. Si immagini una quindicina di metri della riva di
un fiume con una fila ininterrotta di mangrovie con lucciole su ogni foglia
che lampeggino in sincronia, gli insetti sugli alberi ai due estremi della
fila all’unisono con quelli centrali. A quel punto, se si ha
un’immaginazione abbastanza vivida, si puň avere un’idea di questo
sorprendente spettacolo.
Anche piccoli tendenze di anticipare il lampeggiamento successivo con
stimolazioni luminose possono essere sufficienti a creare un ingorgo. Inoltre
in genere non si osserva una propagazione di onde in tale popolazione, tranne
forse quando il lampeggiamento sia appena cominciato o alla fine.
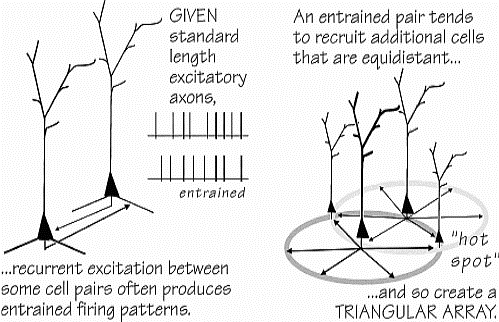
creando
insiemi triangolari equidistanti
|
|
Una
coppia di cellule trascinatetende a reclutare altre cellule
equidistanti,
|
|
eccitazioni
ricorrenti tra gruppi di cellule producono spesso modelli di
accensione trascinati.
|
|
Dati assoni di lunghezza standard,
|
|
Oscillatori a rilassamento come
i neuroni e le lucciole entreranno in sincronia molto piů rapidamente degli
oscillatori armonici, e saranno sufficienti anche interconnessioni deboli
(Somers e Koppell, 1992), cosicché, se parecchi neuroni a distanza di 0,5 mm
sono accesi per qualche ragione (magari perché rispondono al colore giallo),
c’č un’alta probabilitŕ che saranno occasionalmente in sincronia.
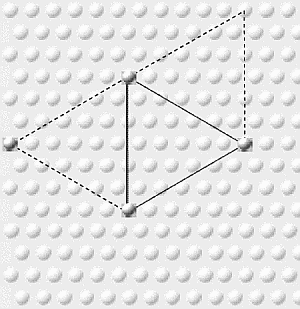 Tale
fenomeno determina una conseguenza interessante: il trascinamento di ulteriori
neuroni che rispondono al colore giallo posti
a 0,5 mm da ciascuno degli originari gruppi di due, a formare un triangolo
equilatero di neuroni piramidali superficiali sincronizzati, come si puň
notare se si osserva la corteccia dall’alto. Naturalmente gli stessi neuroni
del giallo 2 e del giallo 3 potrebbero trascinare un quarto. Ne deriva un meccanismo
per la formazione di un insieme triangolare di cellule sincronizzate, che
estenda il proprio raggio d’azione ovunque ci siano cellule giŕ accese, o
prossime all’accensione. Supponiamo
ora di avere altre cellule che rispondano a un'altra caratteristica di una
banana gialla, ad esempio la sua curvatura: anche esse potrebbero formare il
proprio insieme triangolare. Per capire quale possa essere il piů alto numero
di insiemi triangolari, si cerchi di definire una zona che contenga solo un
membro di ogni insieme triangolare: avrŕ una forma esagonale (i punti
corrispondenti in esagoni adiacenti sono connessi da triangoli equilateri),
sarŕ larga 0,5 mm e conterrŕ alcune centinaia di minicolonne. La maggior
parte degli insiemi potenziali sarŕ naturalmente silente; probabilmente meno
di una dozzina č effettivamente accesa, ma anche gli insiemi silenti
potrebbero essere importanti, se infatti si attivano, potrebbero
“annebbiare” il caratteristico modello di accensione spazio-temporale.
Tale
fenomeno determina una conseguenza interessante: il trascinamento di ulteriori
neuroni che rispondono al colore giallo posti
a 0,5 mm da ciascuno degli originari gruppi di due, a formare un triangolo
equilatero di neuroni piramidali superficiali sincronizzati, come si puň
notare se si osserva la corteccia dall’alto. Naturalmente gli stessi neuroni
del giallo 2 e del giallo 3 potrebbero trascinare un quarto. Ne deriva un meccanismo
per la formazione di un insieme triangolare di cellule sincronizzate, che
estenda il proprio raggio d’azione ovunque ci siano cellule giŕ accese, o
prossime all’accensione. Supponiamo
ora di avere altre cellule che rispondano a un'altra caratteristica di una
banana gialla, ad esempio la sua curvatura: anche esse potrebbero formare il
proprio insieme triangolare. Per capire quale possa essere il piů alto numero
di insiemi triangolari, si cerchi di definire una zona che contenga solo un
membro di ogni insieme triangolare: avrŕ una forma esagonale (i punti
corrispondenti in esagoni adiacenti sono connessi da triangoli equilateri),
sarŕ larga 0,5 mm e conterrŕ alcune centinaia di minicolonne. La maggior
parte degli insiemi potenziali sarŕ naturalmente silente; probabilmente meno
di una dozzina č effettivamente accesa, ma anche gli insiemi silenti
potrebbero essere importanti, se infatti si attivano, potrebbero
“annebbiare” il caratteristico modello di accensione spazio-temporale.
Nella
melodia , composta da 5 note, qui rappresentata, accensioni
simultanee corrispondono ad
accordi.
|
|
Abbiamo
quindi un mosaico esagonale, formato dagli insiemi eccitati e da quelli non
attivati da alcuno stimolo; gli esagoni adiacenti hanno modelli di accensione
quasi identici. Gli insiemi della curvatura
e quelli del giallo non devono essere necessariamente sincronizzati tra loro,
come dice una ben nota formula della teoria di legame; essi verranno
probabilmente sostituiti nel tempo. L’accensione spazio-temporale
nell’esagono č a questo punto minima: contiene un membro di ogni insieme ma
mai un doppione. I microcircuiti nervosi dell’esagono di 0,5 mm possono
entrare in risonanza con il modello spazio-temporale. Un modello imposto
potrebbe essere memorizzato mediante potenziamento e depressione a lungo
termine; infatti si puň immaginare di ricostituire il modello
spazio-temporale, se si č interrotto, grazie all’alterazione prolungata
delle forze sinaptiche, rispetto ai ritmi imposti, creando un nuovo bacino di
attrazione e conformando ogni modello spazio-temporale che sia piů o meno
idoneo a quello memorizzato come standard.
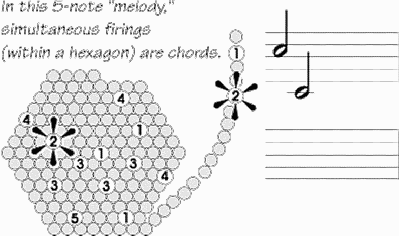
Si tratta di un assemblaggio cellulare Hebbiano minimo, potenzialmente
capace di registrare le diverse caratteristiche di un oggetto o i dettagli di
una serie di movimenti, e il modello di accensione spazio-temporale di un
esagono č potenzialmente un codice cerebrale, ciň che rappresenta un oggetto
o un idea. Se si fa corrispondere a ognuna delle varie centinaia di
minicolonne una nota musicale, tale modello č come un motivetto; si avranno
motivi musicali diversi per caratterizzare una Mela
o una Banana.
La similitudine con la musica spiega inoltre che un mosaico esagonale č
come un coro piano nelle ultime fila di un coro gregoriano. Quando gli insiemi
triangolari reclutano membri ai margini, esagoni addizionali si aggiungono al
mosaico: forse scegliere tra una mela e una banana per merenda dipende da
quanto grandi siano i rispettivi cori, e solo quelli maggiori hanno accesso ai
mezzi espressivi associati al linguaggio del silenzio.
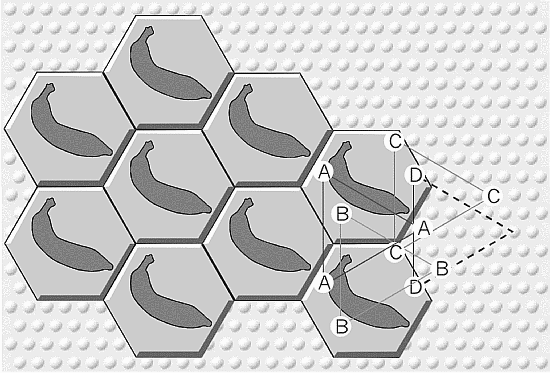
Cori
in competizione per il concetto
Si
immaginino cori in competizione adiacenti a mosaici esagonali che cantino
motivi musicali diversi, nel tentativo di reclutare membri a discapito
dell’altro. Al confine tra i due campi di battaglia ci sono esagoni con
entrambi i motivi sovrapposti, come in un’opera sinfonica. Se tale
combinazione risuona bene con la rete neuronale locale, si parla di armonia,
esattamente come per le scale musicali maggiori e minori. Sovrapposizioni di
confine, cosě come quelle piů estese che possono essere create da fasci
corticocorticali, illustrano un potente principio di ricombinazione, un modo
di creare ricordi associativi che rappresentano relazioni, impiegando lo
stesso codice esagonale di distanza a 0,5 mm usato per gli oggetti.
 Un altro esempio di livelli consiste nel fatto che meccanismi
sufficienti a un livello possano avere fondamenta poco stabili, e che
alternative per lo stesso risultato siano maggiormente estendibili. I codici
esagonali rappresentano fondamenta molto piů solide per superstrutture come
la codificazione di analogie, che sono gli esempi meglio conosciuti di
meccanismi di memoria associativa a livello di meccanismi sinaptici per
condizionamento classico.
Un altro esempio di livelli consiste nel fatto che meccanismi
sufficienti a un livello possano avere fondamenta poco stabili, e che
alternative per lo stesso risultato siano maggiormente estendibili. I codici
esagonali rappresentano fondamenta molto piů solide per superstrutture come
la codificazione di analogie, che sono gli esempi meglio conosciuti di
meccanismi di memoria associativa a livello di meccanismi sinaptici per
condizionamento classico.

Anche quando le risonanze per Mela,
Banana e Ciliegia
sono tutte omnipresenti, un modello puň riuscire a clonare con
maggiore successo, a causa di influenze estrinseche, che
arrivano da altre regioni del cervello. Nella figura si puň
osservare le risonanze per Mela inglobare quelle per Banana
e Ciliegia.
|
|
La
connessione che genera modelli spazio-temporali deve poter
produrre, per condizioni inziali diverse, dozzine di
“melodie” attive, proprio come il midollo spinale genera
modelli spazio-temporali per i movimenti.
|
|
Non č questo il luogo per mostrare le varie
implicazioni di uno schema di codificazione cerebrale basato sull’accensione
spazio-temporale in uno dei cosiddetti esagoni a eccitazione ricorrente, un
progetto la cui trattazione richiederebbe un libro intero e di cui mi sono
occupato in The Cerebral Code. Grazie alla nozione di mosaici
esagonali in competizione transitoria per lo spazio nella corteccia
associativa, si puň comprendere il funzionamento del processo darwiniano
nella corteccia associativa, attraverso modelli spazio-temporali che si
copiano a vicenda lateralmente.
Come
negli esempi classici di un collaudato processo darwiniano, un modello
viene copiato (infatti ciň che č copiato fedelmente definisce
il modello spazio-temporale di forma esagonale), si verificano variazioni
(diseccitazioni, nodi sfocati, sovrapposizioni), le popolazioni dei modelli
varianti competono per uno spazio di
elaborazione, il loro successo relativo č influenzato da un ambiente
(sensore attuale o risonanze con modelli memorizzati), il modello attuale
che ha maggiore successo tende a produrre un numero maggiore del ciclo
successivo di varianti; il principio
dell’ereditarietŕ di Darwin č implementato poiché mosaici piů grandi
hanno un perimetro maggiore, e il perimetro č laddove le diseccitazioni e i
nodi sfocati possono sfuggire alla standardizzazione imposta dai sei nodi
attorno, tutti accesi contemporaneamente.
Diversamente dai tempi generazionali dei normali esempi darwiniani che
si misurano dai giorni ai decenni,
la corteccia opera su una scala temporale che va dai millesimi di secondo ai
secondi, anche se le sue funzioni sono influenzate dai ricordi, che si
misurano secondo scale temporali molto piů lunghe. In un arco di tempo
compreso tra secondi e minuti, la neocorteccia deve essere in grado di
implementare tutti i mezzi classici di accelerazione del ritmo evolutivo (ricombinazione
sistematica, parcellizzazione, rapido “cambiamento climatico” e
riempimento delle nicchie vuote).
Una
rete di competizioni darwiniane
A causa della propria
struttura suddivisa e delle connessioni corticocorticali tra le regioni, la
corteccia non si limita alle produzioni darwiniane standard; puň infatti
usare alcuni elementi addizionali, come un processo darwiniano di supervisione,
che influenzi l’attivitŕ di altri processi darwiniani.
Eppure non deve essere un
supervisore eccezionale e particolarmente intelligente; finché non sarŕ
identificato chiaramente qualcosa di piů complesso, l’ipotesi piů ovvia
sarŕ che qualsiasi processo regolatorio sia essenzialmente stupido, forse si
tratta solo di fenomeni caotici su scala maggiore o minore.
Infatti ci sono situazioni che potrebbero abilitare tale evoluzione
interattiva a due livelli, come il ruolo della corteccia frontale orbitale nel
monitoraggio di progressi rispetto a una serie fi funzioni da svolgere, una
meta-sequenza che sembra procedere su una scala diversa da quella di singoli
pensieri e frasi. Non c’č ragione perché le variazioni darwiniane debbano
essere davvero casuali; un lento processo darwiniano potrebbe influenzare o
deviare la direzione generale delle varianti di un processo darwiniano piů
veloce che gestica problemi di livello inferiore, come la percezione e il
movimento su una scala temporale di secondi. Ci potrebbe essere una cascata o
una rete di tali processi darwiniani che operano su livelli diversi nella
gamma di attributi/metafore, o su diverse scale temporali dai millesimi di
secondo alle ore.
La teoria della “macchina darwiniana” neocorticale sembrerebbe
essere al corretto livello di spiegazione della coscienza; non si trova nelle
sinapsi o nel citoscheletro, ma a livello della dinamica che coinvolge decine
di migliaia di neuroni, i quali generano modelli spazio-temporali, che
rappresentano i precursori di schemi comportamentali nel mondo esterno al
cervello.
Codici
cerebrali compositi, formati da sovrapposizioni e sviluppati da meccanismi di
competizione darwiniana nei processi di copiatura, potrebbero spiegare molto
della vita mentale subconscia. I codici stessi sono adeguatamente arbitrari,
come enfatizzato da un secolo di dibattiti sui simboli. I meccanismi di
competizioni nei processi di copiatura suggeriscono perché gli esseri umani
hanno molti piů comportamenti originali degli altri animali (vantiamo
un’evoluzione “offline” di movimenti non standard, giudicati per
sicurezza), indicando che siamo in grado di impegnarci in ragionamenti
analogici (le stesse relazioni sentimentali possono avere codici che competono).
Poiché i codici cerebrali si formano da singole parti, si puň immaginare un
unicorno e crearne un ricordo (le risonanze che riattivano il codice
spazio-temporale di unicorno). Un processo darwiniano fornisce soprattutto una
macchina per la metafora, cosě da poter codificare le relazioni tra altre
relazioni, e poi modellarle in qualcosa di qualitŕ.
Niente di tutto questo emerge naturalmente dalla “coscienza quantica”,
ma non si faccia di un’erba un fascio: alcune proposte di campi quantici
presentano aspetti interessanti.
Quanti
rivisitati
A proposito della nozione
di sincronizzazione per porre in relazione i diversi aspetti dell’analisi di
un oggetto nel cervello, c’č chi ha parlato di un’influenza in sincronia
di campi quantici, suggerendo una connessione tra i Qm e il riconscimento
dell’oggetto, e combinando contemporaneamente percezione e coscienza. Credo
che i miei insiemi triangolari sincronizzati per la clonazione di un modello
dimostrino un’altra possibile applicazione dello stesso ragionamento,
estendendo il campo potenziale dei Qm oltre gli aspetti della coscienza
relativi al legame oggetto-caratteristica, fino al campo delle funzioni
superiori.
Se č possibile che i fenomeni a livello quantico influenzino alcuni
aspetti della coscienza incoraggiando la sincronia, č vero perň che ipotesi
straordinarie (Qm che facciano da mediatori per un aspetto chiave della
coscienza) richiedono prove straordinarie. Se i fisici della coscienza fossero
seri riguardo alle proprie proposte, esaminerebbero le tantissime soluzioni
alternative per ottenere un sincronismo cerebrale e illustrerebbero perché la
propria spiegazione d sincronizzazione fosse migliore di spiegazioni piů
semplici che sono piů vicine alle funzioni superiori di almeno una dozzina di
livelli.
Un
avvertimento perň: se annunciassi un’altra soluzione per
ottenere una sincronizzazione corticale attraverso qualche nuovo meccanismo
sinaptico, susciterei noia generale; considerando quanti modi di incoraggiare
il trascinamento esistano, dato che anche l’inibizione ricorrente puň
creare trascinamento (si veda piů in generale Winfree, 1967), il nome di
questo “gioco” dovrebbe essere “meccanismi probabili”,
piů che “meccanismi
possibili”. Č importante allora chiedersi quanto sia forte l’influenza di
sincronizzazione del proprio candidato, se altri meccanismi noti sembrino
interagire con esso, e infine se tale candidato sia utile;
sono convinto della necessitŕ di un processo darwiniano di bootstrapping
della qualitŕ, ma alcuni di noi non credono che gli attributi dell’oggetto
abbiano tipicamente bisogno di sincronismo per legarsi insieme, in seguito a
divergenze su dove e cosa. L’esistenza di altri meccanismi di
sincronizzazione indica che ogni effetto dei Qm potrebbe facilmente passare
inosservato, quindi chi proponga questa tesi deve dimostrare non solo
l’effetto dei Qm sul trascinamento, ma anche le condizioni in cui tale
effetto puň prevalere sugli altri, e mettere cosě in luce la “coscienza
quantica”.
Questo fatto mi ha portato a chiedermi se le proposte che si basano
sulla meccanica dei quanti non siano soluzioni in cerca di un problema. Non
sono convinto che la neurologia abbia bisogno di meccanismi basati sulla
meccanica dei quanti quando abbiamo giŕ cosě tanti candidati a livello di
proprietŕ emergenti di circuiti nervosi per ottenere lo stesso scopo, cioč
indurre il trascinamento tra i neuroni corticali, lasciando perdere il
passaggio dal sincronismo ai legami, dai legami all’attenzione,
dall’attenzione ad altri aspetti della coscienza.
Ma questa č scienza, e la tecnologia č un’altra questione, dato
l’altro mio intento riguardo al trascinamento, quello cioč di fornire le
basi per un mosaico esagonale che possa impegnarsi in una competizione
darwiniana. Incoraggiare il trascinamento, o spezzarlo, attraverso campi
applicati dall’esterno, potrebbe offrire un punto d’incontro con i
processi cerebrali darwiniani che spostano i contenuti della coscienza. Se gli
effetti quantici postulati si dimostrano abbastanza forti, e i campi esterni
possono essere focalizzati abbastanza chiaramente da essere portati a
prevalere su altre influenze di trascinamento, il “faretto” rappresentato
dai Qm potrebbe essere alla base di un’interessante
tecnologia. In psichiatria, si potrebbero
 interrompere
i pensieri ossessivi o
interrompere
i pensieri ossessivi o
Treni di pensiero
come
strati di sashimi che sbiadiscono
|
|
eliminare le allucinazioni; in neurologia si potrebbero
riabilitare le zone corticali allo svolgimento di una funzione, persa in
un’altra regione in seguito a un tumore o a un ictus; in anestesiologia, si
potrebbero spezzare quei cori che determinano la memorizzazione di eventi
spiacevoli. Studiare per gli esami con un “aiuto quantico” potrebbe
contribuire all’apprendimento di nuovo materiale; focalizzare tale aiuto in
altre aree potrebbe migliorare la capacitŕ di ricordare.
Flusso
di coscienza
Le risonanze sono oggi meglio conosciute come parte
della serie di attrattori caotici. Si potrebbe trattare di una rete neurale di
esagoni che supportano un numero di modelli spazio-temporali caratteristici,
proprio come i circuiti nervosi del midollo
spinale supportano andature diverse, il particolare modello spazio-temporale
che si ottiene a seconda di come si precondizionino le connessioni nervose
attraverso agevolazioni da parte di altri modelli imposti. Tale ipotesi
potrebbe spiegare qualcosa a proposito del “flusso di “coscienza”.
La manipolazione della zona di un bacino di attrazione richiama alla
mente i treni di pensiero di William James, quella serie di stati mentali che
precedono quello attuale, ciascuno dei quali sbiadisce nel sottofondo, pur
giacendo sui precedenti, e tutti in grado di contribuire a quelle connessioni
che si potrebbero fare anche in questo
momento.
Si paragonino i numerosi attrattori che sbiadiscono a quella tecnica
giapponese che consiste nel tagliare a fette sottili alcune qualitŕ di pesce
crudo, per poi ribaltare il
blocco di lato (un altra analogia, se non si ha familiaritŕ con il sashimi,
č quella con i pezzi del domino). Lo strato inferiore č forse il piů
difficile da raggiungere, ma č anche il piů profondo. Uno scenario
caratterizzato da strati multipli di schemi che sbiadiscono puň contribuire a
promuovere la creativitŕ, mettendo gli strati di attrattori appropriati in un
ordine approssimativamente corretto e adeguando quindi le loro forze relative;
la Teoria Sashimi della Creativitŕ
sarebbe certo un successore adeguatamente “fresco” a tutti quegli schemi
“mal cucinati” sull’emisfero destro del cevello.
Tali digressioni possono risultare perň fuorvianti, e spesso si cerca
di lasciarle svanire, evitando di provocarle con ulteriori ragionamenti. Ci
sono ovviamente diverse tecniche per liberare la mente: Donald Michael (dichiarazione
personale, 1995) suggerisce che la meditazione che impiega un mantra potrebbe
proprio consistere nel creare regioni formate da grandi esagoni semi-statici,
eliminando le preoccupazioni quotidiane che altrimenti smembrerebbero lo
spazio di elaborazione e creerebbero nuovi attrattori a breve termine. Chi
medita, sostituendo tutti i propri pensieri con la formula priva di senso del
mantra, continuando a recitarla abbastanza a lungo da consentire ad
alterazioni di forza sinaptica temporanee di ritornare gradualmente alla
normalitŕ, ottiene un nuovo punto di partenza, per attivitŕ diverse dal
mantra stesso.
Un normale mantra certamente non “svuoterŕ” la mente del tutto:
per cancellare prematuramente quegli attrattori che sbiadiscono, si avrŕ
bisogno di un mantra piů complesso che interrompa le connessioni. A parte
l’annebbiamento con crisi convulsive delle terapie di elettroshock,
personalmente non conosco nessuno schema di eliminazione, sebbene si possano
immaginare virus cerebrali (Dawkins, 1993), che svuotano l’ingresso in quei
bacini di attrazione che sbiadiscono, piů simili a uno strato di vernice
coprente che a una cimosa vera e propria.
Aspetti superiori della coscienza
Una volta concluso con gli aspetti elementari quali
trasformazioni percettive e ricordi, le teorie sulle funzioni cerebrali devono
spiegare astrazioni e associazioni diverse, quali categorie, idee astratte,
schemi, testi scritti, sintassi e metafore. Se si procede oltre la nozione
elementare di coscienza in quanto mera consapevolezza o spostamento di
attenzione, si devono tenere in considerazione i seguenti punti:
o
come
vengono rappresentati i segni linguistici;
o
come
vengono conservati e riportati alla memoria i ricordi;
o
come
si determina il processo di sviluppo darwiniano;
o
come
nascono “nuove idee”,
magari come varianti o sovrapposizioni;
I suddetti quattro punti fanno parte
a grandi linee della ricerca per gli elementi essenziali darwiniani. I
successivi sono piů complessi.
o
L’esistenza
di allucinazioni e sogni.
o
Le
esperienze di déjŕ vu.
La clonazione, molto comune, di un input (talvolta a causa di assenza di
competizione) puň creare l’esperienza cosciente, spesso associata a forti
risonanze mnemoniche, che consentono la clonazione diffusa.
o
Falsi
ricordi.
Tale fenomeno puň essere molto comune, poiché le connessioni sinaptiche
possono essere modificate da un nuovo modello attivo. Allo stesso modo, si
vorrebbe una spiegazione appropriata per il ragionamento concreto e l’idea fissa.
o
Come
sono connesse le varie accezioni della parola “pesca”, per esempio, dato che sono probabilmente conservate
in zone corticali diverse.
o
La presenza di regioni corticali specializzate che possano anche
partecipare ad attivitŕ non specialistiche.
o
La
capacitŕ di mantenere un particolare comportamento, dopo aver scelto fra diverse alternative. Come si
presenta un programma di compiti da svolgere?
o
Funzioni
specializzate di ordine seriale deputate al linguaggio e ai programmi
speculativi,
per non parlare di tutte le sequenze di piccoli movimenti muscolari di cui un
bambino ha bisogno per allacciarsi le scarpe.
o
Connessioni
alla velocitŕ del pensiero, meccanismi che potrebbero variare di volta in volta. Durante le
transizioni caratteristiche della sindrome maniaco-depressiva, un individuo puň
alternare fluiditŕ nel creare connessioni e prendere decisioni a un lento ed
elaborato treno di pensiero che esita troppo a lungo e non riesce a creare
connessioni ovvie.
o
L’impressione
di un narratore, che manipola decisioni e specula sul futuro.
Ogni spiegazione deve essere coerente con l’evidenza neurologica che nessuna
lesione corticale parziale elimina l’ “io”.
Tale č il genere di copertura necessaria a un’utile teoria della
mente, e questa “macchina darwininana” neocorticale permette di effettuare
previsioni su tutto lo spettro (Calvin, 1996). Non č strettamente necessario
che essa spieghi due secoli di neurologia, uno di psicologia e mezzo di
neurobiologia e neuroscienze cognitive, ma non puň essere completamente
incoerente con essi. Una teoria della mente deve avere un grande potere
esplicativo ed essere al contempo abbastanza specifica da effettuare
previsioni sperimentali.
Finora si č avuto bisogno di due metafore: una dall’alto verso il
basso (top-down), che mappa i pensieri su insiemi di neuroni, e una dal basso
verso l’alto (bottom up) che si occupa di come le idee emergano da quegli
insiemi neuronali apparentementi caotici. La “macchina darwininana”
neocorticale puň sostituire entrambe le metafore: in realtŕ č piů un
meccanismo che una metafora, grazie a quei treni espresso nella corteccia.
Passerň ora a trattare l’auto organizzazione di modelli complessi,
utilizzando tali competizioni darwiniane per imprimere risonanze adeguate nel
“feltro” neurale con l’acquisizione di esperienza.
La
ricerca di modelli nascosti
La passione per la ricerca di modelli sembra dipendere fortemente dalla
nozione di coscienza. Se si vogliono avere esperienze significative e
correlate, quelle che siamo in grado di comprendere e su cui possiamo
ragionare, si devono saper discernere i modelli di azioni, percezioni e
concezioni. Dietro la vasta rete di significati letterali interconnessi (tutte
quelle parole su oggetti e azioni) ci sono quelle immaginarie strutture di
comprensione, come lo schema e la metafora, e come l’immaginazione, che ci
permette di estrapolare un percorso, o zumare in avanti su una parte del tutto,
o ancora zumare all’indietro finché gli alberi si fondono, a formare una
foresta.
La prima infanzia implica numerose sfide nella scoperta di modelli, e i
bambini sembrano imparare facilmente modelli sempre piů complessi, nascosti
nei suoni e negli eventi che si determinano attorno a loro. Nel primo anno di
vita, si scoprono69 i fonemi contenuti nelle parole; dopo un altro
anno di apprendimento verbale, si č impegnati a scoprire schemi e sintassi, i
quali costituiscono i legami tra le parole, successivamente si scoprono catene
di subordinate e principi narrativi, nell’ambito di discorsi piů ampi. Le
sovrapposizioni esagonali, cosě come le diverse voci di un’opera sinfonica,
dimostrano come nuove associazioni possano essere rappresentate nel cervello e
l’aspetto darwiniano suggerisce come la qualitŕ potrebbe procedere
attraverso le normali fasi di variazione, competizione e ereditarietŕ.
Da adulti, quando pensiamo seriamente, lo facciamo in maniera ancora piů
astratta. Evochiamo immagini semplificate della realtŕ che chiamiamo concetti
o modelli; possiamo anche scoprire modelli in situazioni speculative, come
quando creiamo un ragionamento deduttivo, utile soprattuto quando si parla di
coscienza! Come scrisse Paul Valéry, il pensiero č tutto ciň che concerne
“quel che non esite, quel che non mi si mostra, quel che fu e quel che sarŕ,
il possibile e l’impossibile”.
La consapevolezza passiva, cosě come i suoi correlati neurali, puň
essere molto piů semplice dei costrutti creativi implicati dai livelli di
coscienza di James, Piaget e Popper; una rapida identificazione di
un oggetto consueto puň non aver bisogno di un meccanismo di competizione per
il processo di clonazione con alternative, come invece una percezione ambigua
o un movimento originale. Certo nel cervello non ci sono soltanto mosaici
esagonali ; infatti essi rappresentano probabilmente solo un modo di
operare di alcune aree neocorticali, e sono regolati da altre regioni
cerebrali, quali l’ippocampo e il talamo. Tali mosaici temporanei sembrano
comunque essere in grado di spiegare molti aspetti della mente, aspetti di cui
č difficile immaginare lo sviluppo a partire dalla meccanica dei quanti,
dalla chimica, dai neurotrasmettitori, da singoli neuroni, semplici circuiti
nervosi, o persino da moduli neocorticali piů piccoli, come le minicolonne.
In alcune regioni, le competizioni esagonali potrebbero essere occasionalmente
l’evento principale.
Ecco quindi emergere da
questa visione del cervello umano, con il suo inflessibile riassetto che varia
da un momento all’altro, alcune idee sulle basi neurali su cui costruiamo il
nostro modo di esprimerci e sviluppiamo i pensieri, alcune possibilitŕ per
implementare il linguaggio e il pensiero razionale. I cori in competizione
sono a un livello di spiegazione che sembrerebbe appropriato al processo
creativo e a quello decisionale. Sarŕ interessante osservare in che misura lo
stesso processo di bootstrapping della qualitŕ, scoperto da Darwin nel 1838,
sarŕ in grado di fornire una spiegazione sulla coscienza interiore.