William H. Calvin University of Washington Box 351800 Seattle WA 98195-1800 USA Email || Home Page || publication list |
as it appeared in copyright ©1975 by authors and publisher. |
William H. Calvin University of Washington Box 351800 Seattle WA 98195-1800 USA Email || Home Page || publication list |
as it appeared in copyright ©1975 by authors and publisher. |
Brain Research, 83 (1975) 498-503 Elsevier Scientific Publishing Company, Amsterdam -Printed in The NetherlandsDouble spikes during otherwise rhythmic discharge were frequently observed in 'fast' pyramidal tract neurons in response to steady depolarizing currents injected through the recording micropipette. The extra spike appears to arise from a large depolarizing afterpotential crossing the falling threshold several milliseconds following a spike; extra spikes themselves may generate further extra spikes in a similar manner, generating burst patterns which are strikingly similar to those of epileptic neurons.
Cerebral cortex neurons with extra spikes:Department of Neurological Surgery,
a normal substrate for epileptic discharges?WILLIAM H. CALVIN and GEORGE W. SYPERT
University of Washington School of Medicine,
Seattle, Wash. 98195 (U.S.A.)(Accepted October 14th, 1974)
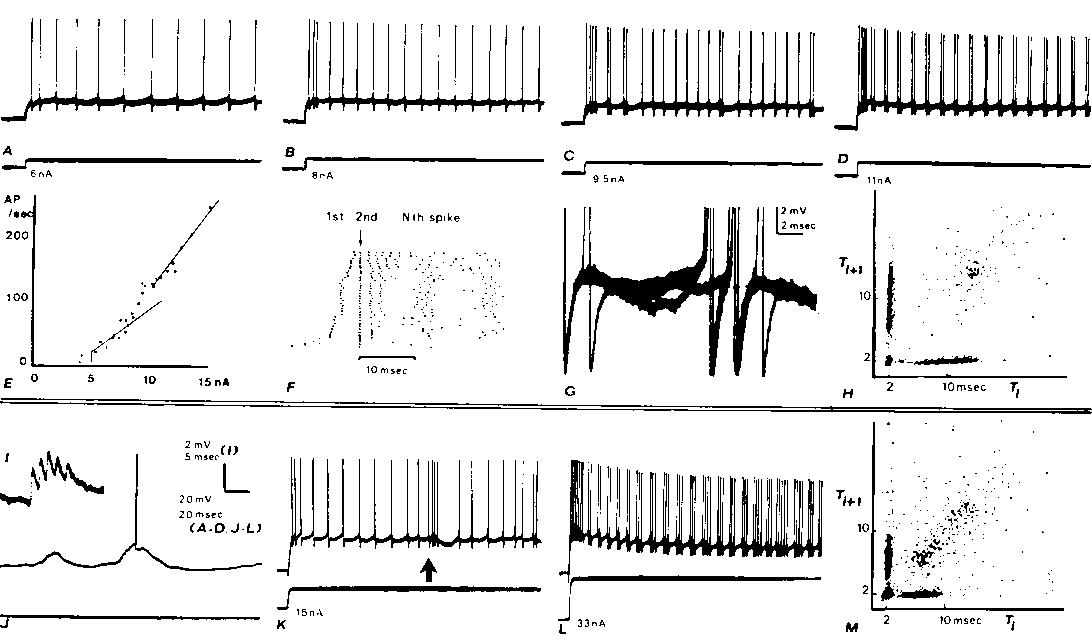
Fig. 1. Intracellular recordings from two cortical neurons (A-H, I-M) with axons projecting into the medullary pyramids with fast conduction velocities. Both neurons exhibit doublet discharges during otherwise rhythmic firing from steps of current injected through the recording micropipette (A). Rhythmic firing evoked by a 6 nA step. B: in response to 8 nA, the rhythmic firing rate increases and a 2 msec interspike interval is seen after the second rhythmic spike. C: a doublet discharge is seen occasionally during otherwise rhythmic firing at 9.5 nA. D: with 11 nA (and greater) currents, doublet discharges recur rhythmically. E: plot of adapted firing frequency versus the injected current. At low currents before doublets are seen during adapted discharge, the frequency-current relation has a slope of 13.6 spikes/sec/nA. At higher currents, when doublet discharge is well-established during maintained firing, the slope is 25 spikes/sec/nA. The intermediate points which have not been fitted with straight lines are those currents at which mixed responses were elicited (as in C). F: raster displays which aligns the second spike of each response below the arrow: dots representing the first spikes appear to the left of the arrow. Data from many different current steps (range as in E) arranged in ascending order. The 11 nA step was repeated 12 times. G: membrane potential trajectory between spikes during adapted firing; 3 superimposed sweeps triggering upon a spike several hundred msec after the beginning of the train in C. In 2 cases, no doublet occurred and a hump can be seen following the spike; in the third case, an extra spike is seen arising from a level just above this hump. H: joint interval density is a scatter plot of interspike interval versus the following interspike interval. Plot mixes the responses from the entrire range of current values shown in E and F. The 'arrow pointing to the origin' shape of this plot is characteristic of the fast PT neurons which develop doublets at higher rhythmic rates (see M). I-M: similar records from another fast PT neuron. The effects of a large synaptic input associated with a spindle in the electrocorticogram (ECoG) are shown in I-K. I: large unitary-appearing events in synaptic noise. I: spindle associated synaptic waves arising from the �80 mV resting potential elicit occasional spikes. K: low-frequency rhythmic response to a current step is interrupted by a spindle wave at arrow; note gradual rise in firing rate. L: after exhibiting a progressive development of doublets as illustrated for the other neuron in A-D, this neuron developed triplets and even longer clusters of extra spikes at higher currents. (Baseline changes are due to tip potential alteration with current.) Thus high-frequency bursts may be due to either large synaptic waves (as in K) or to an extra spike mode of rhythmic firing. M: joint interval density as in H.
Supported by NIH Grants NS 04053, NS 05211 and NS 09677. We thank Jerrold Maddocks and Susan Johnston for their technical assistance.1 ATKINSON, J. R., AND WARD, A. A., JR., Intracellular studies of cortical neurons in chronic epileptogenic foci in the monkey, Exp. Neurol., 10 (1964) 285-295.